Жаба озерна
? Жаба озерна | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||
Охоронний статус | ||||||||||||||||||
 Найменший ризик (МСОП 3.1) | ||||||||||||||||||
Біологічна класифікація | ||||||||||||||||||
| ||||||||||||||||||
Біноміальна назва | ||||||||||||||||||
Rana ridibunda Pallas, 1771 | ||||||||||||||||||
 | ||||||||||||||||||
Посилання | ||||||||||||||||||
|
Жаба озерна (Pelophylax ridibundus, синонім Rana ridibunda) — вид земноводних, що входять до підроду зелених (водяних) жаб (Pelophylax Fitzinger), роду Зелена жаба (Pelophylax), родини Жаб'ячі (Ranidae) ряду безхвостих земноводних (Anura).
В європейській частині в озерної жаби підвидів не виділяють.
Вид, як вже зазначалося, відомий тим, що бере участь в утворенні гібридогенних комплексів (на території України такий комплекс утворений озерною, ставковою та їстівною жабами). У цій групі озерна жаба, схрещуючись з P. lessonae, формує окремий таксон P. esculentus, існування якого, як правило, підтримується шляхом гібридизації з одним з батьківських видів, хоча відомі популяції, представлені одним гібридогенним таксоном. При цьому утворення статевих клітин в особин гібридного таксона пов'язано з вибірковим збереженням генома лише озерної жаби (неменделівське спадкування). Разом з тим останнім часом виявлені популяції, специфіка спадкування в яких не відповідає цій схемі.
Зміст
1 Поширення
2 Чисельність
3 Зовнішній вигляд
4 Мінливість
5 Генетика
6 Життя у природі
7 Охорона
8 Посилання
Поширення |
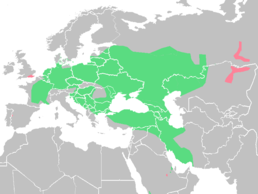
Озерна жаба живе на величезній території, що простягається у широтному напрямку від Східної Франції до Казахстану. Встановлення точних меж видового ареалу доволі складне у зв'язку зі змінами думок про таксономічний статус окремих популяцій та популяційних груп і, крім того, у зв'язку з сучасною інтродукцією цих амфібій у ряді регіонів. Тим не менше можна стверджувати, що у Західній Європі в межах своєї ділянки ареалу цей вид на півночі доходить до Балтійського узбережжя, у східній — до північної Латвії (околиці Риги, інтродуценти), північного заходу Росії (околиці С.-Петербурга) та крайнього півдня Фінляндії, Удмуртії, Башкирії (Уфа), і далі в Азії — до північно-східного Казахстану. На півдні — до північного Ірану, Афганістану, Пакистану, північно-західного Китаю.
Ізольовані популяції відомі на заході Саудівської Аравії (Азир), Бахрейні, Аль Хазі та в деяких інших регіонах Аравійський півострів|Аравійського півострова. На крайньому заході за межами свого ареалу інтродуковані популяції виявлені в Іспанії, на південному заході Франції та на
крайньому сході — в околицях Якутська.
В окремих випадках дата першого випуску в природу та основні етапи розселення відомі. Так, у Східному Казахстані у 1961 та 1974-75 рр. Р. ridibundus випуще ні в Усть-Каменогорську, згодом виявлені в Алакольській котловині біля селища Уч-Арал (1967 г.), потім в околицях Семипалатинська та Павлодара (80-і роки), ще згодом — у північно-західній частині Зайсанської котловини (1998). На Середньому Уралі (Єкатеринбурзька обл.) озерна жаба інтродукована у 1969 р. у Верхньотагільському водосховищі з Краснодарського краю, а в 1980 р. — у Рефтинське водосховище з Одеської області. У центрі Горно-Алтайська виявлені у 1970 р. (озерних жаб постачали до місцевого педінституту з Ошської обл., Киргизстан), а через 14 років ці амфібії заселили більшість водойм в околицях міста.
У викопному вигляді виявлена у нижньому плейстоцені-голоцені середньої течії Дніпра.
В Україні озерна жаба водиться практично по всій території у місцях, де є прісні водойми. У виборі водойм віддає перевагу біотопам відкритих просторів.
Враховуючи те, що антропогенні зміни торкаються практично більшої частини території країни (включно із зведенням лісів), можна вважати, що Р. ridibundus живе у всіх природних зонах України. Разом з тим у своєму поширенні у гірських районах Закарпаття вона звичайно не піднімається вище 600 м і 1150 м — в Криму.
Спектр біотопів, що їх займає озерна жаба, доволі широкий і включає місця, які обов'язково характеризуються наявністю постійних прісних водойм: береги річок, озер, ставів, стариць, дренажні канави, водойми-відстійники тощо. На півдні України (Присивашшя, північно-східна частина степового Криму та Керченського півострова) озерні жаби можуть займати водойми з підвищеною твердістю та солоністю.
Чисельність |
Загалом можна зазначити, що детальні дослідження динаміки чисельності озерних жаб та факторів, що зумовлюють її мінливість у різних біотопах як на території України, так і за її межами, обмежені. Наявні дані говорять про істотну мінливість чисельності цих тварин у різних ділянках ареалу та в різних біотопах.
Так, на Закарпатті їхня чисельність становить 15-45 ос. на 100 м маршруту. При розрахунку числа особин на одиницю площі цей показник досягає 1 ос. на 0,3-2 м² водойми, при цьому на Прикарпатті щільність цього виду дещо менша й коливається в межах 1 ос. на 7,5-40,0 м².
На території Криму чисельність Р. ridibundus у різних ділянках коливається від поодиноких особин на 1 км маршруту до декількох десятків або навіть сотень.
Кількість озерних жаб у Кривому Розі та його околицях (р. Інгул та р. Саксагань) на 10 м² берегу досягає 30-50 ос., у міських водоймах до 10-16, на каналі Дніпро — Кривий Ріг до 50 ос. На одному кілометрі каналу Дніпро — Донбас на початку 90-х років інкубувалося понад 82 млн ікринок. При цьому чисельність пуголовків, що успішно завершили метаморфоз, становила близько 0,3 млн.
Із схожих даних, одержаних при вивченні цих амфібій за межами України, варто згадати те, що в гирлі Ками (до створення Куйбишевського водосховища) відзначалось по 5-6 озерних жаб (з них 1-2 самиці) на 2-4 м² берегу, у Пестречинському районі Татарстану — до 30 особин на 200 м² нерестової водойми (Гаранин, 1983). Загалом на території Волзько-Камського краю Росії чисельність цих амфібій коливалася в межах 0,1-28,0 ос. на 100 м берегової смуги, хоча безпосередньо на річках та ставках вона істотно більша й досягала 50,6-97,5 ос.
Судячи з оглядових матеріалів С. Кузьмина (1999), чисельність Р. ridibundus у південних регіонах Росії становить близько 170—1410 ос./км берегової лінії, а в долині
Амудар'ї в Туркменистані приблизно 180—860 ос./км.
Доволі висока чисельність амфібій цього виду й у Верхньотагільському водосховищі Свердловської області (інтродуковані з Краснодарського краю) — до 1500 ос./км.
Зовнішній вигляд |

Тварини середніх та великих розмірів, довжина тіла яких може досягати понад 100 (до 170) мм, у забарвленні спинної частини тулуба переважають різні відтінки зеленого кольору (інколи до коричневого), черевний бік тіла звичайно світлий з неясно виявленими невеликими сірими плямами, які часто створюють мозаїчномармурове тло (у Криму виявлені популяції озерних жаб з білим забарвленням черевного боку та браком плям).
Характерною особливістю візерунка є розвиток світлої спинної поздовжньої смуги (яка може заходити на голову) та наявність округлих темних плям, розміри яких коливаються від великих (досягаючи приблизно розмірів ока), до середніх та дрібних. По боках морди в області очей може бути розташована темна смужка, що інколи може переходити на тулуб (у пониззі Дніпра виявлені популяції з розвитком сіро-брунатної смуги в області барабанної перетинки, що нагадує таку в бурих жаб).
На верхній частині передніх (передрамено) та задніх (гомілка та заплесноплеснева частина) кінцівок як правило, розвинені темні поперечні смуги. Є спинно-бокові складки. Позад очей містяться добре виявлені барабанні перетинки. Якщо стегна розташувати перпендикулярно серединній осі тіла, то надступаковогомілкові (скакові) зчленування заходять одне за одне. Пальці на задніх кінцівках з'єднані плавальними перетинками, внутрішній п'ятковий горбок невисокий.
Самці звичайно відрізняються від самиць розвитком на першому пальці передніх кінцівок темно-сірих шлюбних мозолів та наявністю в кутах рота темносірих звукових резонаторів.
Мінливість |

Пуголовок жаби озерної
Озерна жаба, порівняно з іншими видами, найчастіше ставала об'єктом досліджень зовнішньої морфології безхвостих амфібій. Одна з перших основоположних праць у цьому плані належить П. В. Терентьєву (1943). Він показав, що в розвитку кінцівок та деяких параметрів очей існує зв'язок зі статтю, а відносна довжина задніх кінцівок, відношення L. o./L. c., Sp. ip./Lt. p., T./F. та L. t. ci./D. h. мало залежать від віку.
Перші узагальнення оригінальних даних з мінливості зовнішньої морфології озерних жаб України зроблені В. І. Таращуком (1959), М. М. Щербаком (1966), М. М. Щербаком та М. І. Щербанем (1980). Так, деякі розмірні показники, без уточнення точок збору наведені В. І. Таращуком, який відзначає, що L. озерних жаб з території України звичайно не перевищує 10-11 см, Lt. p. більше у 1,5 (і більше) разу від Sp. ip., L. tym. вкладається у L. o. 1,23-1,60 разу, F. / T. 0,87-1,01, L. t. ci. / D. h. відповідає 2,0-4,5.
М. М. Щербак вказує, що за 13 стандартними розмірними показниками озерні жаби Криму істотно відрізняються від тварин цього виду з Молдови (11 статистично достовірних відмін). Крім того, наведені також результати порівняння кримських Р. ridibundus з особинами з віддаленіших ділянок ареалу — Кавказу та Киргизстану. І хоча зроблено формальний висновок про поширення у всіх трьох регіонах номінативної форми, його обґрунтованість ускладнена тим, що жаби у східних ділянках ареалу могли належити, як стало відомо багато пізніше, до різних таксонів. З другого боку, озерним жабам Криму властива, найбільш вірогідно, географічна мінливість. У всякому разі, судячи з останніх порівнянь передгірно-гірських та степових популяцій, між ними існують достовірні відміни за L. o., D. n.-o., D. r.-n. та Sp. in.
За зовнішньоморфологічними показниками статистично достовірні відмінності між Р. ridibundus Прикарпаття та Закарпаття існують лише за довжиною стегна. Дещо східнішим популяціям амфібій цього виду з Київської та Черкаської областей властива залежність від статі за індексами F. / T., D. p. / L. t. ci., T. / L. t. ci. та Ix (Ix — мультиплікативний індекс, що розраховується як T2 ?D.h./L.t.ci.2 ?L. c. s.;). Цікаво, що Ix згідно з розрахунками склав тут не менше 32, що істотно відрізняється від цього показника, вказаного його автором для цього виду раніше — не менше 45.
Пропоноване пояснення різниці у розмірах цього показника географічною мінливістю ніяк не можна вважати обґрунтованим у зв'язку з браком відомостей про точне походження матеріалу в праці С. В. Таращука (1985).
Особливу цікавість викликає відміна між календарним (визначається за шаруватістю кісткової тканини) та біологічним віком амфібій. У популяціях з центральної України представлені 3 розмірно-вікові групи: напівдорослі (самиці 66,1, самці 66,8; тут і далі розміри в мм), дорослі-1 (самиці 81,8, самці 80,1) та дорослі 2 (самиці 92,2, самці 83,5).
Із збільшенням віку виявленість статевого диморфізму посилюється, а деякі з ознак, що традиційно використовуються в таксономії зелених жаб, у представників цих трьох вікових груп можуть достовірно відрізнятися.
Порівняння зовнішньоморфологічних ознак озерних жаб українських популяцій з аналогічними показниками представників цього виду із східних частин ареалу і в тому числі з Центрально-Чорноземного р-ну (Росія), Удмуртської республіки (Росія), Татарстану (Росія), Вірменії та Киргистану вказує на відмінності, пов'язані, скоріше за все, з географічною мінливістю.
У західних ділянках ареалу (Нідерланди, Польща, Румунія) порогові значення індексу T./L. t. ci. (L. t. ci.=C. int.) Р. ridibundus досягають 8,5-9,5 (Pagano et al., 1995).
Аналіз особливостей виявлення візерунка та забарвлення цих амфібій у природних популяціях поки що також не дає можливості виявити будь-які виразні закономірності просторової мінливості цих ознак, хоча специфіка спадкування деяких з них вже відома: наявність смуги контролюється домінантним алелем, а брак — рецесивним станом гена.
Озерним жабам із Закарпатської, Одеської, Херсонської, Донецької областей та з Криму властива відсутність статевих відмінностей у розвитку спинної смуги та характері плямистості верхньої та нижньої частин тіла. Для 58,5% особин з цього регіону характерний брак спинної смуги, а в 42,3% вона є (з них в 11,5% носить переривчастий характер), у 77,7% розвинені великі (близько 6-8 мм в діаметрі) темні спинні плями, у 56,2 і 76,9% плямисте черево та горло відповідно.
Однак у жаб з передгірно-гірської частини Криму плямистість черевного боку тіла відсутня більш ніж у 96% особин, а з степової зони — у 64-65%, хоча відомо, що для
більшості озерних жаб характерний мармурово-плямистий візерунок черева. Разом з тим в Одеській області у межах однієї вікової групи смугасті особини частіше трапляються серед самців, ніж серед самиць (92,9 і 76,2% відповідно). При цьому серед смугастих частіше трапляються особини з асиметрією плям, однак відмін між самцями та самицями немає. У всіх чотирьох вибірках виявлена позитивна кореляція (коеф. Спірмена, рівень значущості порядку 0,05) асиметрії плям з наявністю смуги.
Згідно з аналізом понад 400 особин з території України (точні місця зборів невідомі), наявність спинної смуги також характерніша для самців (56%), і саме в них частіше розвинена плямистість на черевному боці тіла (20,5% проти 14,7%). У плямистості спинного боку статевий диморфізм не виявлений: ця ознака властива 94% самців і 99% самиць.
Більшості Р. ridibundus (86,4%) з околиць Києва характерне зелене чи зеленобрунатне забарвлення тла спинного боку тіла. Що ж до наявності світлої спинної смуги, то частота її трапляння тут становить 80,4%. Смуга може мати пряму або зиґзаґувату форму (у 52,7% і 27,7% відповідно). Майже всі Р. ridibundus (понад 95%) характеризуються пігментацією горла та черва та наявністю на передніх кінцівках 3 і приблизно 10 смуг на задніх кінцівках.
Привертають до себе увагу особливості мінливості цих ознак P. ridibundus за межами України. Так, в озерних жаб (Краснодарський край, Росія та Абхазія) забарвлення черевного боку тіла корелює з віком. Серед дорослих тварин (довжина тіла більше 61 мм) краснодарської популяції чисельність темнозабарвлених (наявність різних варіантів пігментації горла, черева та стегон) становить 40,6%, а серед молодих довжина тіла менше 60 мм — лише 5,3%, тобто коефіцієнт кореляції r досягає 0,75 (p<0,001).
Разом з тим, у другій вибірці (ст. Ірклієвська) зв'язок забарвлення черевного боку тіла з віком відсутній. Особливо слід підкреслити сезонну мінливість частоти трапляння цієї ознаки, що виявлялося в обох вибірках. Так, у вибірці з околиць Краснодара навесні частота світлозабарвлених становить 44,1%, темнозабарвлених 19,3% (в окол. ст. Ірклієвська 44,4 і 9,5% відповідно), восени у першій вибірці світлозабарвлених 9,6% проти 38,1% темнозабарвлених (в окол. ст. Ірклієвська 13,9 проти 23,4% відповідно). Понадто, відзначена більша частота трапляння смугастих жаб навесні (49,0%) порівняно з осінню (26,0%).
Цікавість викликає інформація про те, що в популяціях P. ridibundus у Липецькій області Росії тварини з плямистістю спинного боку не виявлені (Статкевич, 2001).Ще далі на північний схід (м. Єкатеринбург) серед цьоголітків частота трапляння жаб без медіальної смуги становить 56,6%, а серед дорослих — відсутня повністю.
На території Татарстану частота трапляння смугастих особин не залежить від статі й коливається у межах 65-70%, але зі збільшенням розмірів особин цей показник зменшується з 80 до 50%.
Далі на схід (Середній Урал, Свердловська обл.) в інтродукованій популяції озерної жаби морфа striata (розвиток медіальної смуги) у 1976 р. складала 66%, а в 1981 р.— 35,1%. Особливу цікавість викликає той факт, що за умов експерименту в цьоголітків, вирощених за підвищених температур, частота трапляння цієї ознаки зростає з 15,9% до 29,8%.
У Волгоградській області чисельність особин з розвиненою спинною смугою переважає над безсмугими. Так, частота трапляння в одній популяції смугастих самців та самиць складає відповідно 74,9-71,1%, в іншій 68,2-71,4% (відмінності за статтю не достовірні в обох вибірках). Частота тварин із забарвленим горлом та черевом у першій популяції практично не відрізняється (57,4 і 57,0 самці й самиці відповідно), у другій відміни складають близько 12% (самці — 75,9%, самиці — 64,3%).
Серед інших морфологічних особливостей цих тварин можна вказати на розвиток різного роду аномалій. Так, в індустріальних районах Дніпропетровської області кількість тварин цього виду з різними відхиленнями в будові кінцівок коливається від 34,3% до 47,6% (у 1991—1994 рр. досліджено 3505 особин).
Порівняння мінливості середньої частоти асиметричного прояву на ознаку залежно від висоти місцевості (використовували тварин лише з незабруднених водойм) показало, що на висоті близько 1000 м цей показник коливається в межах 0,35-0,36, а на рівнині він дорівнює 0,47. У Р. ridibundus у зонах з різним ступенем антропогенного навантаження (у міській смузі Казані та за її межами) середня частота асиметричного прояву на ознаку дорівнює 0,72-0,75 і 0,35 відповідно.
У Самарській області у місцях з послабленим антропогенним навантаженням цей показник коливається в межах 0,50-0,51, а на забруднених ділянках 0,55-0,66. Ще один показник ступеня забрудненості територій відбивається у частоті трапляння клітин з порушеннями: у незабруднених ділянках в межах цієї області вона становить 1,5-3,0%, а в забруднених — 5,33%. В Астраханській області у контрольній (незабрудненій) ділянці середня частота асиметричного прояву на ознаку дорівнює 0,49, а в зонах з підвищеним антропогенним впливом — від 0,60 до 0,67 (результати мікроядерного тесту: 1,15 та 1,68-3,00% відповідно).
Цікаво, що у Казахстані (Семипалатинська обл.), розміри тіла особин із забруднених ставів перевищують такі в жаб з незабруднених водойм на 5%, а маса — на 12%, індекс L. c. / L.— на 12%. Морфологічні аномалії в цих амфібій на ділянках з різним антропогенним навантаженням у Казані відрізняються. У зоні з сильним промисловим забрудненням частота особин з подвоєнням першого пальця на задніх кінцівках коливається від 0 до 8,7%, а в зеленій зоні 0-3,3%, при цьому відзначені також й інші відхилення.
Біоакустичний аналіз шлюбних криків зелених жаб з території колишньої Югославії, вказує на видоспецифічність цього показника в представників усіх таксонів цієї групи (P. ridibundus, Р. lessonae, Р. esculentus). При порівнянні шлюбних криків (тривалість, частота, кількість імпульсів) озерної жаби з представниками інших таксонів цього комплексу (Франція) виявилося, що P. ridibundus відрізняється від двох останніх представників, які на дендрограмі показують більшу близькість між собою, ніж с озерною жабою.
Вивчення мінливості цього показника в даних тварин на території Ізраїлю показало його залежність від температури навколишнього середовища. Іншим фактором, що впливає на специфіку цього показника, є розміри тварини. Для більшості параметрів шлюбних криків P. ridibundus також властива залежність від температури, за винятком впливу останньої на групові пульси (вони в середньому становлять 3,55; популяції з території колишньої Югославії).
Генетика |
Каріотип P. ridibundus на території України містить 26 хромосом (2n = 26, NF = 58) та дещо відрізняється від інших видів цієї групи особливостями морфології хромосом (значення центромерного індексу); на обох хромосомах 9-ї пари локалізовані вторинні перетяги.
Каріотип озерної жаби зі східної частини Німеччини також складається з 26 хромосом (2n = 26,NF = 58), але вторинні перетяги відзначені на 10-й парі хромосом.
Хромосомні набори озерних жаб з території Кабардино-Балкарії (Нальчик), Грузії (Поті) та України (Львів) відповідають каріотипам озерних жаб з інших частин ареалу (2n = 26, NF = 52), при цьому вторинні перетяги наявні на 11-й парі і, крім того, існує тенденція інших хромосом також утворювати вторинні перетяги.
Вищевказані відмінності у вказівках про локалізацію вторинних перетягів на різних парах хромосом (на 9-й, 10-й або 11-й) скоріше за все пов'язані з різними методичними підходами дослідників до ідентифікації гомологів. Хромосомні набори амфібій цього виду з території Московської області Росії принципово не відрізняються від наведених вище даних.
Особливу цікавість викликає дослідження метафазних хромосом та хромосом типу лампових щіток за допомогою С-забарвлення (Bucci et al., 1990). Озерні жаби відрізняються від інших членів групи виразно розрізнюваними темними гранулами та флуоресцентним ефектом у районі центромерів. Специфічність також виявляється в особливостях центромерів, кількості та локалізації гігантських петель, а також у частоті хіазм.
В одних і тих самих особин можуть бути відмінності в кількості хромосом. Крім звичайного для цього виду диплоїдного набору 2n = 26 у клітинах кісткового мозку, в клітинах сім'яників нараховують 26, 39, 52 та 78 хромосом (Манило, 2005). Крім поліплоїдних клітин (близько 10%), також відмічаються близько 5% клітин з анеуплоїдною кількістю хромосом (матеріали дослідження потребують уточнення та їхньої інтерпретації).
Кількість ядерної ДНК в Р. ridibundus з Одеської області та Криму становить 14,2-14,6 пг; використання іншого методу для визначення цього показника в озерних жаб у східних областях (Харківська, Луганська та Донецька) показало, що розмір генома досягає в середньому 16,12 пг, коливаючись у межах 15,74-16,35 пг. В інших частинах ареалу (Росія) цей показник змінюється незначно.
Популяції озерної жаби право- та лівобережної України характеризуються високим рівнем їх гетерозиготності (Н = 0,05-0,087) при відносно невеликій генетичній віддаленості одного від одного (DN = 0,151 і DN = 0,129 відповідно); при цьому слід вказати, що рівень гетерозиготності їх дещо перевищує середній для всіх амфібій (Не = 0,067) і коливається в межах H0 = 0,05-0,087.
Життя у природі |
Амфібії цього виду виходять з місць зимівлі залежно від погодних умов весни кожного року, але найчастіше у березні-квітні. Понадто, у теплі зими на півдні України (південний берег Криму) озерні жаби можуть бути активними цілорічно. Це справедливо й для тих водойм у дещо північніших ділянках ареалу, що пов'язані з теплими водами накопичувачів електростанцій, скидним ділянками очисних споруд, артезіанськими свердловинами, що функціонують цілорічно тощо. Парування починається незабаром після виходу з місць зимівлі й зазвичай припадає на квітень (кінець березня у теплі роки), і сезон нересту триває майже до кінця червня-липня.
Особливу цікавість викликають дані з двох інтродукованих популяцій на Середньому Уралі (Свердловська обл.), у яких відзначалась різна тривалість строків відкладання ікри. Так, в інтродукованій популяції Рефтинського водосховища (з Одеської обл., Україна) відкладання ікри починається у травні й тривалість нересту становила всього три тижні.
Озерні жаби у Верхньотагільському водосховищі, узяті з краснодарських популяцій півдня Росії характеризується, тривалістю нересту з кінця квітня до середини липня (Иванова и др., 2001).При цьому частина личинок не встигає завершити метаморфоз і зимує на личинковій стадії, що призводить до мінливості розмірів цьоголітків однієї й тієї самої генерації від 16,7 до 50,0 мм. Є вказівка про те, що ікра відкладається на глибині не менше 0,4-1,5 м (твердження потребує перевірки).
В озерних жаб кримських популяцій кількість ікринок в одній кладці коливається в межах 5-10 тис. У Придніпров'ї (Дніпропетровська обл.) середня абсолютна плодючість Р. ridibundus становить близько 5400 (850-12400) ікринок, а відносна 106 (40-280) ікринок на 1 г маси тіла самиці.
Кількість ікринок в одній кладці залежить від розмірів самиці (за даними з популяцій Краснодарського краю). Так, самиці з розмірами тіла в межах 50,1-60 мм характеризуються величиною кладки в 1,1-4,2 тис. ікринок; 60,1-70,0 мм — від 3,0 до 5,3 тис.; 70,1-80,0 мм — від 3,0 до 5,8 тис.; 80,1-90 мм — від 4,1 до 8,8 тис.; 90,1-100,0 мм — від 7,1 до 16,4 тис. і понад 100 мм—от 4,9 до 9,6 тис. ікринок. Чисельність личинок, що вийшли з ікринок з цих кладок, становить 40,0-68,8%, а кількість тих, що пройшли метаморфоз 0,4-2,2% (по відношенню до кількості відкладеної ікри). З 16 кладок озерних жаб з цього ж регіону, число ікринок від однієї самиці приблизно 4240 штук, а вихід — близько 31,8%.Що ж до чисельності личинок, що пройшли метаморфоз, то вона становить близько 20,8% (або 6,6% по відношенню до кількості відкладеної ікри).
На території Українських Карпат загальні розміри личинок 6-8 мм при виході з яйця, а перед метаморфозом (настає через 70-90 днів), близько 65 мм. Пуголовки озерної жаби з цього регіону характеризуються величиною проміжку між очима приблизно у 2 рази більшими, ніж дистанція між ніздрями. Анальний отвір розташований з правого боку, зябровий — зліва й скерований назад-угору.
У східних частинах ареалу (Караганда, Казахстан) відзначено випадок масового гігантизму личинок, які досягали загальних розмірів близько 150 мм; матеріали схожого дослідження у західних ділянках ареалу (Латвія) показали, що такі личинки можуть характеризуватися розвитком органів, властивих наземній стадії — легенів та статевої системи (Лусис, Цауне, 1984; Aitbaeva et al., 1991).
Порівняння співвідношення статей серед дорослих озерних жаб у кримських популяціях показало незначне переважання кількості самиць (59 проти 41%). Інші дані, щодо статевої і вікової структури українських популяцій озерних жаб практично відсутні. Докладніша інформація з цього питання існує щодо амфібій цього виду в інших частинах ареалу. Так, серед самиць Р. ridibundus 6-9(10)-річного віку, що розмножуються, з території Вірменії близько 20% досягають статевої зрілості вже у 3 роки, а 80% починають розмноження після 3-ї (4-ї) зимівлі (самці стають статевозрілими у віці 2-4 років).
Чисельність самців та самиць озерних жаб на території Волзько-Камського краю (Росія) відразу після метаморфозу співвідноситься як 1 : 2, хоча є відомості, що частка самиць збільшується у старших вікових групах (тобто зі збільшенням розмірів особин) від 40 до 100% (Татарстан). У північніших ділянках ареалу (Росія, Івановська обл.), виявлено, що максимальний вік Р. ridibundus становить 7-11 (12) років, при цьому статевої зрілості жаби досягають в середньому в 4-5 років, а середній вік самиць приблизно на 0,7-1,4 року перевищує вік самців.
Серед інтродукованих на Середньому Уралі (Свердловська обл.) озерних жаб майже не трапляються тварини старші 6 років, більшу частину популяції (до 70%) становлять 3-4-річні особини.
Матеріали з добової активності в Україні обмежені твердженнями про цілодобову активність Р. ridibundus у Карпатах протягом весни — початку літа (Никитенко, 1959), а за іншими спостереженнями — переважанням активності вдень від 12 до 15 години та в присмерку від 20 до 21 години (Щербак, Щербань, 1980). На території Криму озерні жаби рано навесні та восени активні тут протягом дня, а влітку — цілодобово.
Амфібії цього виду досить добре переносять високі температури, і їх неодноразово відзначали за температур повітря й води близько 30-35°С і 25-28°С відповідно. Межі нижніх температур повітря близько 2,5-10°С, води — приблизно 8 °C (дані для кримських популяцій; Щербак, 1966). В інших джерелах (Кузьмин, 1999) наведені інші показники без зазначення конкретного регіону (Центральна Азія?): верхні межі температури повітря до 35-40°С і води близько 30 °C, нижні — 10-18°С повітря і 5-17°С води.
Тривалість личинкового розвитку становить близько 80-90 днів, але істотно залежить від специфіки умов водойми, у якій відбувається розвиток і, передовсім, від її температурного режиму. Наявність у колекціях (Зоологічний музей ННПМ Національної академії наук України) серії личинок з весняними та пізньоосінніми датами зборів (31 березня 1961 р., с. Зеленогір'я та 4 листопада 1961 р., околиці Алушти) дає підстави твердити, що не всі пуголовки встигають пройти метаморфоз і вони зимують на цій стадії.
На зимівлю йдуть звичайно у жовтні-листопаді, й ці строки залежать від особливостей осені кожного року. Зимують у воді.
Пуголовки живляться детритом, водоростями, вищими рослинами та тваринними об'єктами, при цьому різноманітність об'єктів з віком личинок збільшується, але бентосні об'єкти все одно й надалі складають основу живлення.
Аналіз живлення дорослих жаб на Кримському півострові (Щербак, 1966) показав, що найчастіше в їхньому раціоні трапляються личинки двокрилих, павуки, туруни, мурашки, свинки, полукрильці тощо (відповідно 22,3, 21,4, 18,1, 13,6, 11,7%). Спектр живлення амфібій цього виду з території Закарпаття приблизно схожий і тут відзначені мурашки (49,3%), личинки двокрилих та попиків (по 29%), турунів (20,3%), мершавиць (17,4%).
У раціоні неодноразово відзначалися й хребетні (мальки риб, ящірки, пташенята біляводних птахів і т. ін.). У більшості випадків P. ridibundus полюють на берегах водойм у межах 1 метра або трохи більше того від урізу води.
Вивчення живлення цих амфібій в інших частинах ареалу (дельта Волги) показало, що озерні жаби з середини червня припиняють покидати водойми, і основою їхнього живлення у цей період стають пуголовки свого виду, а згодом — цьоголітки (виявлені у 80% шлунків при аналізі спектра живлення 500 особин). З інших кормів виявлені личинки та імаго бабок і великих водяних жуків (23%), 11% молюсків, 8% вовчків та 5% мальків риб.
У деяких місцях (у вузькій зоні біля шлюзу Ільменя), частка риби досягала 50% частоти трапляння від інших об'єктів живлення, але після відкриття шлюзу й відходу мальків, останні зникають із спектру живлення. Крім того, якісний склад об'єктів живлення залежить від багатства окремих видів. Так, у місцях з високою чисельністю вовчків частота їх трапляння складала 93%; у шлунках жаб, спійманих біля вуликів, бджоли зареєстровані майже з 100%-ю частотою; у місцях концентрації молоді сазана вони також відзначені в 100% Р. ridibundus.
Разом з тим важливо підкреслити, що за матеріалами цього дослідження відносна чисельність вилучених амфібіями мальків риб (від загального числа випущених рибгоспом), складала всього близько 0,06-0,08% (за даними аналізу шлунків 1400 особин).
Серед прямих ворогів дорослих озерних жаб відзначені чимало видів біляводних рептилій, птахів та ссавців. Понадто, ці земноводні можуть складати основну частину живлення окремих видів хребетних — вужів (Natrix natrix, N. tesselata), гомілкових птахів (Ciconiiformes), норки (Mustela) та деяких інших.
Попри те, що Р. ridibundus є чи не найлабільнішим видом серед амфібій вітчизняної фауни, поміж основних факторів, що справляють вплив на чисельність популяцій, слід вказати усе ж на антропогенний прес. Одним з прикладів може слугувати випадок прориву греблі, що огороджувала скидання вод Рижовського цукрового заводу в р. Синюху (притока р. Південний Буг) у 1985 р. Обсяг скидних вод склав близько 7000 м3 і при цьому температура води підвищилася до 42 °C, а показник рН досяг рівня 7. Чисельність озерних жаб після цього знизилася до 1-2 ос./100 м берегової лінії (проти 69-128 до скидання вод).
Охорона |
Вид занесено до Додатку III «Бернської конвенції» (категорія «види, що підлягають охороні») та до «Червоної книги хребетних Міжнародного союзу охорони природи (МСОП)».
Посилання |
Писанець Є. Земноводні України (посібник для визначення амфібій України та суміжних країн). — Київ: Вид-во Раєвського, 2007. — 197 с.
| Цю статтю потрібно вікіфікувати, щоб привести її вигляд до стандартів Вікіпедії. (Листопад 2011) |


