Безхвості
? Безхвості Час існування: тріас - сучасність | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Зелена деревна жаба (Litoria caerulea) | ||||||||||||||
Біологічна класифікація | ||||||||||||||
| ||||||||||||||
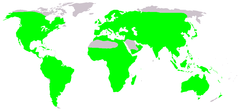 Поширення безхвостих (зеленим) | ||||||||||||||
Підряди і родини | ||||||||||||||
Archaeobatrachia
Mesobatrachia
Neobatrachia
| ||||||||||||||
Посилання | ||||||||||||||
|
Безхво́сті (Anura, Salientia, Ecaudata) — ряд земноводних, який є найбільшим із сучасних, нараховуючи понад 6000[1] відомих науці видів. Личинку безхвостого земноводного називають пуголовком.
Знахідки викопних форм, дуже схожих з жабами, відомі з нижньої юри, однак не всі дослідники включають їх до даного ряду. Найдавніші безперечні представники безхвостих відомі починаючи з середньої юри. Серед найдавніших є Triadobatrachus massinoti. В теперішній час безхвості поширені по всій земній кулі, від тропіків до субарктичних регіонів, найбільше різноманіття видів спостерігається у тропічних дощових лісах.
Розміри дорослих безхвостих варіюють у межах від 7,7 мм (Paedophryne amauensis) до 320 мм (Conraua goliath). Представники ряду мають кремезний тулуб, випуклі очі, роздвоєний язик і зігнуті під тулубом кінцівки; хвіст відсутній (звідки й назва). Середовище проживання безхвостих на всіх стадіях розвитку включає прісні водойми, а для дорослих особин — додатково суходіл, крони дерев і підземні нори. Найбільш характерний спосіб пересування безхвостих — це стрибки, але різні види опанували додаткові способи, такі як ходіння і біг, плавання, лазіння по деревах і планерування.
Покрив безхвостих має багато залоз, шкіряні виділення деяких видів надзвичайно отруйні. Скупчення залоз у деяких видів створюють враження, що шкіра вкрита бородавками, багато інших видів мають гладеньку шкіру. Забарвлення безхвостих варіює від маскувального бурого, зеленуватого і сірого до найяскравіших червоних, жовтих і чорних кольорів, що зазвичай сигналізують про отруйність (або є мімікрією під неї). Шкіра безхвостих проникна для води, але різні адаптації дозволяють їм уникати надмірної втрати вологи при сухопутному способі життя.
Як правило, безхвості відкладають ікру у воді. З ікринок вилуплюються водні личинки, пуголовки, що мають зябра і хвости. Пуголовки проходять стадію метаморфозу, в кінці якої перетворюються у дорослих особин. У той же час, деякі види відкладають ікру на суходолі, а інші не проходять стадію пуголовка. Дорослі особини більшості видів є хижаками, але є й всеїдні види і навіть декілька видів, які харчуються фруктами. Безхвості здатні видавати широкий діапазон звукових сигналів, особливо під час шлюбного сезону. Спостереження за безхвостими виявили складні схеми поведінки у шлюбних ритуалах, при загрозі життю і в інших ситуаціях.
Багато видів безхвостих (за деякими даними, до третини від всіх видів земноводних) перебувають під загрозою вимирання[2].
Люди вживають безхвостих у їжу, утримують як домашніх тварин, крім того, безхвості слугують зручними модельними організмами для біологічних досліджень.
Зміст
1 Еволюція
2 Ареал
2.1 Поширення в Україні
3 Анатомія і фізіологія
3.1 Загальна характеристика
3.2 Опорно-рухова система
3.2.1 Кінцівки
3.3 Шкіра
3.4 Отруйність
3.5 Дихальна система і кровообіг
3.6 Травна система
3.7 Видільна система
3.8 Репродуктивна система
3.9 Нервова система
3.10 Органи зору
3.11 Органи слуху і нюху
3.12 Анабіоз
4 Класифікація
5 Комунікація
6 Пересування
6.1 Стрибання
6.2 Біг і ходіння
6.3 Плавання
6.4 Риття нір
6.5 Лазіння по деревах
6.6 Планерування
7 Самозахист
8 Життєвий цикл
8.1 Розмноження
8.2 Ікра
8.3 Пуголовки
8.4 Метаморфоз
8.5 Дорослі особини
8.6 Піклування про потомство
9 Використання людиною
9.1 Утримання в неволі
9.2 В кулінарії
9.3 В науково-дослідній сфері
9.4 Виробництво отрути
10 Охоронний статус
11 Безхвості в культурі
11.1 У фольклорі
11.2 В літературі
12 Примітки
13 Література
Еволюція |

Скам'янілість жаби з Чехії, можливо, Palaeobatrachus gigas
Походження безхвостих і їх еволюційні взаємозв'язки з іншими сучасними земноводними залишаються предметом дебатів[3][4].
Prosalirus bitis — одна з найдавніших тварин, що має багато ознак безхвостих. Скам'янілі рештки цього виду були вперше виявлені у 1995 році в Аризоні, у нижньоюрських відкладеннях формації Кайєнта (англ. Kayenta Formation) віком понад 180 мільйонів років[5]. Prosalirus ще не мав потужних задніх кінцівок, але його посадка (структура таза) нагадує сучасних жаб, він не мав хвоста[6][7] і був адаптований до пересування стрибками[8]. Інша ранньоюрська жаба описана під назвою Vieraella herbsti. Цей вид відомий лише за одиничною знахідкою — відтиском черевної і спинної сторони однієї особини. Припускають, що її розмір становив 33 мм.
Еволюційні лінії Prosalirus bitis, Vieraella herbsti та деяких інших викопних видів відокремились ще до того, як з'явився найближчий спільний предок сучасних жаб. Тому деякі автори не включають дані види до складу ряду безхвостих[9]. Найдавнішим відомим науці представником безхвостих у такому разі виявляється середньоюрський вид Eodiscoglossus oxoniensis, близький сучасним Bombinatoridae і Alytidae[3].
Викопні безхвості були виявлені на всіх континентах, за винятком Антарктиди, але наявні непрямі ознаки, що вказують не те, що коли клімат на Землі був теплішим безхвості мешкали й там[10].
Ареал |
Безхвості поширені практично по всьому суходолу. Винятками є великі піщані пустелі Сахара і Руб-Ель-Халі, найбільш холодні області — Гренландія, Таймир та інші високоширотні райони Арктики, Антарктида, а також Камчатський край і деякі острови, віддалені від континентів[11][12]. Крім того, природний ареал безхвостих раніше не включав острів Нової Зеландії, але невдовзі після спроб штучного заселення принаймні два види (Litoria raniformis і Litoria ewingii) утворили на ньому стійкі популяції[13]. Багато видів мають обмежене поширення через кліматичні і географічні бар'єри, наприклад, протоки, гірські хребти, пустелі; популяції можуть бути ізольовані також через перешкоди, створені людиною — автотраси, лісні просіки тощо[14]. В тропічних регіонах видове розмаїття, як правило, більше, ніж в районах з помірним кліматом[15]. Деякі види безхвостих адаптовані для виживання у негостинних умовах, наприклад, в пустелях[16] або холодному кліматі. Наприклад, Rana sylvatica, чий ареал частково лежить за північним полярним колом, на зиму закопується в землю. Незважаючи на глибоке промерзання ґрунту, висока концентрація глюкози в тканинах дозволяє цій жабі пережити зиму у стані анабіозу[17].
Поширення в Україні |
Фауна України налічує 13 видів безхвостих земноводних. Найбільш різноманітно представлений рід бурих жаб — це жаби трав'яна, гостроморда (болотяна), прудка, озерна, ставкова та їстівна. Поширені три види ропух: ропуха звичайна мешкає у лісах, садах і луках, у Кримських горах; ропуха зелена — по всій Україні; ропуха очеретяна — у західних областях. Зустрічаються два види кумок: кумка червоночерева — по всій території України, кумка жовточерева — в західних областях. Також в Україні поширені райка деревна з роду райок і часничниця звичайна з роду часничниць.
Анатомія і фізіологія |
Загальна характеристика |

Рентґенівський знімок жаби Paedophryne swiftorum
У дорослих амфібій є парні п'ятипалі кінцівки, типові для наземних хребетних тварин. Тіло широке, коротке і сплощене. Дорослі особин не мають хвоста (що й визначило назву ряду); хвостовий відділ хребта видозмінений у паличкоподібну кістку уростиль; ребра відсутні; хвіст втрачають під час метаморфозу. Мають добре розвинені кінцівки; задні кінцівки довші ніж передні, мають міцнішу мускулатуру і звичайно пристосовані до стрибання. Гола шкіра безхвостих проникна для води і газів, має багато залоз[18].

Скелет їстівної жаби. Видно кістки черепа, хребет і пояси кінцівок.
Опорно-рухова система |
Скелет безхвостих, як і у інших наземних хребетних, включає череп, хребет, пояси кінцівок і скелет вільних кінцівок[19]. Череп сплощений, переважно хрящовий[20]. Бічні потиличні кістки утворюють парні потиличні виростки, що з'єднують череп з шийним хребцем.
Хребет безхвостих складається з чотирьох відділів — шийного, тулубного, крижового і хвостового. Шийний і крижовий включають лише по одному хребцю. У тулубному відділі 7 хребців. Всі хребці хвостового відділу зрослись в одну кістку — уростиль[20].
Пояс передніх кінцівок складається з парних лопаток, ключиць та коракоїдів і непарної грудини. Пояс задніх кінцівок складається з клубових і сідничних кісток. Сідничні кістки з'єднані лобковим хрящем[20].
Кінцівки |

Litoria tyleri. Добре видно широкі пальці і перетинки між ними
Будова кінцівок безхвостих у різних видів варіюється і залежить від середовища проживання — наземного, водного чи деревного. Як правило, безхвості здатні робити різкі рухи, завдяки чому вони ловлять здобич і тікають від хижаків. Ефективні способи пересування забезпечуються декількома особливостями будови кінцівок:
- У багатьох безхвостих, які живуть у водному середовищі, наявні перетинки між пальцями. Існує кореляція між відносною площею перетинок і часткою часу, яку тварина перебуває у воді[17]. Наприклад, у африканської жаби роду Hymenochirus, спосіб життя якої виключно водний, плавальні перетинки закривають більшу частину проміжку між пальцями, натомість у австралійської жаби Litoria caerulea, яка проводить більшу частину життя на деревах, перетинки покривають від чверті до половини площі цих проміжків.
- У безхвостих з деревним способом життя, часто можна спостерігати на ступнях присоски, завдяки яким вони тримаються на вертикальних поверхнях. Присоски працюють по принципу капіляра наступним чином: тканина присосок складається з клітин, які здатні під тиском зменшуватись в об'ємі. Коли особина ступає на поверхню дерева, клітини присосок стискаються, залишаючи вузькі канали, наповнені слизом, який утримує тварину на поверхні.
- У багатьох видів деревних безхвостих можна виявити на кожному пальці невеликі подушечки, які збільшують площу стику з деревом. Також багато видів мають характерну особливість будови стегнового суглобу, яка дозволяє їм пересуватися на тільки стрибками (як реша безхвостих), але також ходою. У деревних безхвостих, як живуть на великій висоті, також можна виявити перетинки між пальцями. У цих видів перетинки адаптовані для уповільнення падіння, а у деяких видів навіть для планерування[21].
- У наземних видів вищезгадані адаптації відсутні. Їх задні кінцівки, як правило, мають розвиненішу мускулатуру, у порівнянні з водними безхвостими. У деяких наземних видів, які риють ґрунт, на кінчиках пальців наявні невеликі мозолі, пристосовані для риття.
Розвиток кінцівок може бути ускладнений у деяких випадках:
- Одна з зародкових задніх кінцівок може бути з'їдена хижаком, наприклад, личинкою бабки. У більшості випадків це перешкоджає регенерації повноцінної кінцівки, іноді вона може виявитись понівеченою чи зовсім не розвинутись (тим не менш, тварина може вижити і з трьома кінцівками).
- Паразитичний плоский черв (Ribeiroia trematodes), проникаючи у задню частину тіла пуголовка, може змінити положення клітин зародкових кінцівок. Іноді це призводить до розвитку однієї або двох додаткових кінцівок[22].
Шкіра |

Шкіра жаби під мікроскопом
У багатьох видів безхвостих шкіра проникна для води (найбільш проникна ділянка — область очей). Ця особливість створює для безхвостих загрозу втрати рідини і пересихання. У деяких безхвостих, що ведуть деревний спосіб життя, присутня адаптація у вигляді додаткового водонепроникного шару шкіри. Інші безхвості мінімізують втрату води адаптаціями у поведінці: нічним способом життя, прибиранням поз, що зменшують площу стикання з повітрям[23], наприклад, вони відпочивають у групах і тісно притискаються один до одного[23].

Забарвлення сумчастої ропухи дозволяє їй зливатися з облетілим листям
Шкіра безхвостих допомагає їм маскуватися. Представники деяких видів здатні змінювати відтінок шкіри, щоб краще зливатися з навколишнім фоном[24]. Шкіра безхвостих, що ведуть нічний спосіб життя, часто покрита бородавками і зморшками (що дозволяє їм маскуватися під елементи ґрунту), тоді як у безхвостих, що ведуть деревний спосіб життя, шкіра, як правило, гладенька (ці види частіше мімікрують під елементи рослин)[24].
Деякі види безхвостих змінюють забарвлення в залежності від рівня освітлення і вологості навколишнього середовища. Цю здатність забезпечують спеціальні клітини, наповнені пігментом, розмір яких змінюється під впливом світла і вологості. Зміна забарвлення шкіри від світлішого до більш темного сприяє терморегуляції цих видів[25].
Отруйність |

Отруту жаби Oophaga pumilio використовували для виготовлення отруйних стріл
Багато безхвостих виробляють токсичні речовини, які слугують як для захисту від хижаків, так і для нападу. Наприклад, у ропух отруйні залози розташовані у верхній частині голови над очима[26]. Хімічний склад отрути безхвостих залежить від виду і може містити подразники шкіри, галюциногени, нервово-паралітичні токсини, вазоконстриктори, токсини, що викликають конвульсії, та інші. Хижаки, що спеціалізують на певних видах безхвостих, як правило, адаптовані до властивого цим видам типу отрути, але неспеціалізовані тварини, як і людина, можуть серйозно постраждати від контакту з отрутою безхвостих, що в деяких випадках призводить до летального результату[27].
Джерело отруйних речовин у безхвостих також неоднакове. Деякі безхвості генерують токсини самотужки[28], інші використовують токсини, отримані з їжею (частіше за все членистоногих)[29]. Як правило, безхвості сигналізують про свою отруйність за допомогою яскравого, насиченого, «попереджувального» забарвлення. Існують також неотруйні види безхвостих, які мімікрують під отруйні види своїм забарвленням, яке відлякує хижаків[30][31].
Дихальна система і кровообіг |
Шкіра безхвостих проникна для молекул кисню, вуглекислого газу і води. Кровоносні судини, розташовані прямо під шкірою, дозволяють їм дихати при повному зануренні у воду, оскільки кисень проходить з води крізь шкіру прямо в кров. На суходолі безхвості дихають за допомогою легень. У них відсутня система м'язів (черевних, діафрагми і міжреберних), яка забезпечує у ссавців механізм вдих-видих. Замість неї у безхвостих розтягується горло, повітря входить через ніздрі, а потім стиснення м'язів ротової порожнини вганяє повітря до легень[32]. У серпні 2007 року був відкритий вид жаб Barbourula kalimantanensis, що ведуть повністю водний спосіб життя. Це перший відкритий вид безхвостих, що не має легень[33].
Безхвості мають трьохкамерне серце, як і інші тетраподи (за винятком ссавців, птахів і крокодилів, у яких серце чотирикамерне). Насичена киснем кров з легень і збагачена вуглекислим газом кров з тканин надходить в серце через різні передсердя. Спеціальний клапан регулює подачу крові зі шлуночка серця в аорту чи в легеневу артерію, в залежності від типу крові. Цей механізм забезпечує мінімальне змішування крові з підвищеною концентрацією кисню і крові, насиченою вуглекислим газом, і, таким чином, сприяє більш активному обміну речовин[34].
Деякі види безхвостих адаптовані до виживання у воді з пониженою концентрацію кисню. Так, наприклад, жаба Telmatobius culeus, що мешкає у високогірному озері Тітікака, має зморшкувату шкіру зі збільшеною площею поверхні, що сприяє обміну газів. Як правило, ця жаба не використовує свої рудиментарні легені. Спостереження показали, що представники цього виду, перебуваючи на дні озера, час від часу здійснюють ритмічні рухи вверх-вниз, що збільшує течію води навколо них[35].

1 праве передсердя, 2 легені, 3 аорта, 4 ікра, 5 товста кишка, 6 ліве передсердя, 7 шлуночок черця, 8 шлунок, 9 печінка, 10 жовчний міхур, 11 тонкий кишечник, 12 клоака
Травна система |
Зуби безхвостих, так звані педициллярні зуби, розташовані у верхній щелепі[19], з їх допомогою тварина утримує їжу пере тим, як її проковтнути. Ці зуби недостатньо міцні для укусів і піймання жертви. Свою їжу (мух та інших дрібних рухливих тварин) безхвості ловлять липким, роздвоєним язиком. У пасивному стані язик згорнутий у роті. Він прикріплений спереду до щелеп, і жаба може вистрілити ним вперед і повернути його назад з великою швидкістю[17]. У деяких жаб немає язика, і вони засовують їжу у рот передніми кінцівками[17]. У інших очі можуть втягуватись через отвори у черепі, надавлювати на їжу, що знаходиться у роті, і таким чином проштовхують її в горло[17]. Проковтнута їжа проходить через стравохід до шлунку, де оброблюється травними ензимами. Після цього вона надходить у тонкий кишечник, де триває її перетравлення. Секреція соку підшлункової залози і жовчі, яка виробляться в печінці і накопичується у жовчному міхурі, відбувається в тонкому кишечнику. Там також відбувається максимальна абсорбція нутрієнтів в організм. Відходи їжі переходять у товстий кишечник, звідки після абсорбування надлишків води переміщуються в клоаку.
Видільна система |
Видільна система безхвостих схожа за структурою з системою ссавців. В її основі лежать дві нирки, що фільтрують сечовину та інші продукти життєдіяльності з крові. Отриманий фільтрат нирки концентрують у сечі, яка потім проходить через сечовід і накопичується у сечовому міхурі. З сечового міхура продукти життєдіяльності організму потрапляють в клоаку і звідти — назовні[36].
Репродуктивна система |
Репродуктивна система безхвостих, лише за деякими винятками, заснована на зовнішньому заплідненні. у багатьох видів безхвостих самці менші за самиць. У самців є голосові зв'язки, а у багатьох видів і горлові мішки, за допомогою яких вони видають гучне квакання під час шлюбного сезону. Два сім'яника прикріплені до нирок, сперма проходить через нирки, після чого протравляє у сечоводи, а звідти в клоаку. За відсутності пеніса сперма викидається з клоаки одразу на ікру, відкладену самицею під час амплексуса. У самиць є парні яєчники, розташовані біля нирок. Ікринки проходять через парні яйцепроводи назовні. Під час амплексуса хватка самця стимулює відкладання ікри, як правило, у оболонці, що нагадує желе[36].
Нервова система |
Нервова система безхвостих складається з мозку, спинного мозку і нервів. Багато частин головного мозку безхвостих відповідають людському мозку. Мозок складається з двох нюхових долей, двох церебральних хемісфер, шишкоподібного тіла, мозочка і довгастого мозку. Мозочок контролює м'язову координацію і рівновагу, а довгастий мозок керує диханням, травленням та іншими автоматичними функціями організму[36]. Відносні розміри головного мозку безхвостого набагато менші ніж у людини. Вони мають лише 10 черепних нервів і 10 пар спінальних нервів, у порівнянні з ссавцями, птахами і рептиліями, які мають 12 черепних нервів[36]. У безхвостих немає зовнішнього вуха, а барабанні перетинки відкриті назовні. Як і в інших тетраподів, вуха безхвостих включають в себе вестибулярний апарат. Маючи відносно короткого вушного равлика, безхвості використовують для розпізнання звуків електричний апарат (на відміну від механічного у ссавців)[37].
Органи зору |
Органи зору представлені парними очима. Жаби володіють унікальним серед хребетних зоровим апаратом. У ході досліджень було виявлено, що майже 95% інформації йде в середній відділ мозку, яка використовується лише для здійснення рефлексів. Жаби бачать лише рухомі предмети.
Жаби також «фільтрують» рухомі предмети, помічаючи лише округлі. При цьому вони не звертають уваги на предмети, що рухаються ритмічно вправо і вліво.
Органи слуху і нюху |
Органи слуху складаються з середнього вуха (барабанна порожнина) та внутрішнього вуха (барабанна перетинка). Органи нюху мають численні складки слизової оболонки, що збільшує чутливу поверхню органа. Частина нюхового мішка відокремлюється у самостійний орган, характерний лише для наземних хребетних — якобсонів орган.
Анабіоз |
В екстремальних погодних умовах деякі види безхвостих впадають у стан анабіозу і можуть не проявляти ніякої активності протягом декількох місяців. В холодних областях безхвості взимку впадають в сплячку. Деякі наземні безхвості (наприклад, Anaxyrus americanus) риють собі на зиму нору і сплять в ній. Інші види, менш пристосовані до риття, ховаються в ущелинах або закопуються у сухе листя. Водні види (наприклад, жаба-бик), як правило, залягають на дні водойми, частково занурюючись у мул, але зберігаючи доступ до кисню. Їх метаболізм уповільнюється, і вони виживають за рахунок споживання внутрішніх енергетичних резервів. Багато безхвостих можуть вижити після замерзання. Незважаючи на те, що під їх шкірою і у порожнинах тіла утворюються кристали льоду, життєво важливі органи захищені від замерзання завдяки високій концентрації глюкози у тканинах. На вигляд нежива, замерзла жаба може почати дихати і відновити роботу серця, якщо її відігріти[38].
З іншої сторони, Cyclorana alboguttata під час пекучого сухого сезону в Австралії впадає в стан естивації (літньої сплячки), і обходиться без їжі і води 9-10 місяців на рік. Ця жаба заривається у землю і згортається у захисному коконі, який утворює її скинута шкіра. Дослідження показали, що під час естивації метаболізм змінюється так, що ККД мітохондрій збільшується, дозволяючи більш ефективно використовувати обмежену кількість енергетичних ресурсів, доступних жабі у стані сплячки. Намагаючись дати відповідь на питання, чому цей механізм не має широкого поширення серед тварин, дослідники дійшли висновку, що він корисний тільки для холоднокровних тварин, що перебувають у комі тривалий час і чиї енергетичні затрати вкрай низькі, оскільки у них немає необхідності постійно генерувати тепло[39]. Інше дослідження показало, що для задоволення мінімальних енергетичних потреб у жаби атрофується більшість м'язів (за винятком м'язів задніх кінцівок)[40].
Класифікація |
До ряду безхвостих земноводних відносять всіх сучасних жаб і ропух, а також викопні види, які підходять за описом до безхвостих. Морфологічні характеристики дорослого безхвостого включають, серед іншого, 9 або менше передтазових хребців, довгу, направлену вперед клубову кістку, наявність уростиля і відсутність хвоста, вкорочені у порівнянні з задніми передні кінцівки, спаяні разом ліктьова і променева кістки передніх кінцівок, а також великогомілкову і малогомілкову кістки задніх кінцівок, подовжену щиколотку, беззубу нижню щелепу і підшкірні лімфатичні порожнини, розташовані між шкірою і м'язовим шаром[41]. Личинки безхвостих (пуголовки) мають один центральних дихальний отвір і ротові воронки з кератиновими зубчиками[41].
Безхвостих класифікують на три підряди: Archaeobatrachia, який включає чотири родини примітивних жаб; Mesobatrachia, який включає п'ять більш еволюційно просунутих; Neobatrachia — найбільший підряд, до якого входять решта родин сучасних жаб. Neobatrachia, в свою чергу, класифікують на дві надродини: Hyloidea і Ranoidea[42][43].
Деякі види безхвостих формують міжвидові гібриди. Наприклад їстівна жаба являє собою природний гібрид ставкової (Pelophylax lessonae) і озерної (P. ridibundus) жаб[44][45].
Комунікація |

Жаба їстівна. Добре видно горлові мішки по боках рота, що задіяні при кумканні
Для кожного виду безхвостих характерний свій тип квакання. Квакання — це звук, який виробляться при проходженні повітря через гортань. У багатьох видів є додаткові адаптації, що посилюють звук, — горлові мішки, що являють собою шкіряні мембрани, здатні до розтягування і розташовані під горлом чи по боках рота. У деяких представників безхвостих (наприклад, Heleioporus і необатрахусів) немає горлових мішків, але тим не менш вони здатні видавати гучне квакання, оскільки ротова порожнина у них має форму купола і достатньо великий об'єм, щоб виконувати функцію підсилювача. Квакання деяких видів безхвостих чутно далі ніж на кілометр[46]. Види безхвостих, що мешкають переважно вздовж проточної води, часто не мають адаптацій для посилання звуку, оскільки шумний фон робить звукову комунікацію неефективною[47]. Натомість ці види використовують візуальні способи спілкування («семафорять» один одному).
Спостереження показують, що основна доля комунікації безхвостих припадає на приваблювання самиць. Самці можуть квакати поодинці, а можуть створювати хор, коли багато самців збираються у зручних для парування місцях. Самиці багатьох видів безхвостих (наприклад, Polypedates leucomystax) також квакають у відповідь на заклики самців, що, можливо, підвищує локальну репродуктивну активність[48]. Самиці надають перевагу самцям, що видають найбільш гучні й низькі звуки, які вказують на здорового і міцного самця, здатного породжувати найкраще потомство[49].
Окремий клас звуків видають самці чи незацікавлені самиці, яких осідлав інший самець. Це характерне цвірінькання, яке супроводжується вібрацією тіла[50]. Деревні жаби і деякі види наземних безхвостих мають у своєму репертуарі звук-попередження про майбутню зливу, який вони видають при поєднанні певних кліматичних умов[50], а також специфічний звук, що відлякує чужих самців з зайнятої території. Всі ці звуки безхвості видають із зачиненим ротом[50].
Деякі безхвості в стані смертельної небезпеки видають сигнал лиха з відкритим ротом, отримуючи високий пронизливий звук. Як правило, його видає жаба, схоплена хижаком. Іноді дезорієнтований цим криком хижак випускає жабу, а іноді на крик збігаються інші хижаки, відволікаючи того, що зловив жертву, що дозволяє їй вирватися[50].
Пересування |
Безхвості пересуваються різними способами. В залежності від середовища проживання вони стрибають, бігають, ідуть ходою, плавають, риють нори, лазять по деревах і планерують.
Стрибання |

Colostethus flotator у стрибку
Безхвості вважаються найкращими стрибунами серед всіх хребтових (за довжиною стрибка відносно розмірів тіла)[51]. Австралійська жаба Litoria nasuta може стрибати на відстань, яка перевищує довжину її тіла (5.5 см) у більш ніж 50 разів[52]. Прискорення у стрибку може досягати 20 м/с2. Види безхвостих сильно відрізняються один від одного за своїми здібностями у стрибках. Всередині виду існує позитивна кореляція між розміром особини і довжиною стрибка, але відносна довжина стрибка (довжина стрибка, поділена на довжину тіла) при цьому зменшується. Індійська жаба Euphlyctis cyanophlyctis має унікальну спроможність підстрибувати з води з позиції лежачи на поверхні[53]. Крихітні жаби Acris crepitans можуть «перебігати» по поверхні ставка короткими різкими стрибками[54].
Здатність до стрибків у безхвостих обумовлена тим, що більша частина їх опорно-рухового апарату модифікована для стрибків. Гомілка, фібула (?) і ступні поєднані в єдину міцну кістку, також поєднані променева і ліктьова кістки передніх кінцівок (вони гасять інерцію під час приземлення). Подовжені плесна збільшують дожину ноги, що дозволяє жабі відштовхуватись від землі протягом довшого часу, збільшуючи прискорення. Клубова кістка також подовжена і формує з крижовою кісткою рухомий суглоб, який у еволюційно просунутих безхвостих, таких як Ranidae і Hylidae, функціонує подібно додатковій кінцівці, підсилюючи силу стрибка. Хвостові хребці зрослись в уростиль, розташований всередині тазу. Він дозволяє ефективно передавати імпульс стрибка від ніг до тіла[55].
Подібним чином модифікована мускулатура безхвостих. Подібно іншим тваринам, що мають кінцівки, у давніх безхвостих їх рух регулювався парами м'язів — згиначами і розгиначами. У сучасних безхвостих м'язи, що сприяють стрибку, непропорційно розвинені (основні ножні м'язи складають понад 17% від загальної маси жаби), тоді як м'язи, що повертають кінцівки у початкову позицію, майже атрофовані. Уповільнена зйомка стрибка демонструє, що м'язи ніг можуть згинатись пасивно. Спершу вони розтягаються (при тому, що жаба продовжує сидіти), потім вони стискуються і одразу ж розгинаються знову, відправляючи жабу у повітря. Під час стрибка передні кінцівки притиснуті до грудей, а задній витягнуті у повну довжину[56]. У деяких безхвостих (наприклад, Osteopilus septentrionalis і Rana pipiens), максимальне зусилля, яке докладають м'язи під час стрибка, може перевищувати їх теоретичну потужність. Це відбувається за рахунок того, що після згинання і стискання м'язів першого разу отримана енергія переходить до натягнутого сухожилля, яке обертається навколо кістки щиколотки. При другому стисканні це сухожилля вивільнюється подібно катапульті і надає жабі прискорення, якого було б неможливо домогтися одними лише зусиллями м'язів[57]. Схожий механізм був виявлений у деякий коників, в тому числі сарани[58].
Біг і ходіння |
Безхвості з родин Bufonidae, Rhinophrynidae і Microhylidae мають короткі задні кінцівки і пересуваються крокуючи, а не стрибками[59]. Швидке пересування у представників цих родин забезпечують або прискорений рух кінцівок (швидке ходіння), або швидкі короткі стрибки. Метод пересування Gastrophryne olivacea описаний як «комбінація з бігу і коротких стрибків (лише декілька сантиметрі у довжину)»[60]. В одному з експериментів жаба Bufo fowleri була посаджена на бігову доріжку, яка оберталась з різною швидкістю. При замірюванні кількості кисню, яку витрачала жаба, було виявлено, що для цього виду пересування маленькими стрибками на довгі дистанції є енергетично неефективним, але може бути виправдане для швидкого пересування на короткі відстані[61].
Жаба Kassina maculata має короткі і тонкі кінцівки, не пристосовані для стрибання. Ця жаба може швидко бігати, пересуваючи по черзі задні ноги. Уповільнена зйомка показала, що алюр цієї жаби не змінюється в залежності від швидкості бігу (на відміну, наприклад, від коня, який біжить риссю на середніх швидкостях, а на високих переходить на галоп)[62]. Це вид також вміє залазити на дерева та кущах, чим активно користується уночі для ловлі комах[63]. Індійська жаба Euphlyctis cyanophlyctis має широкі ступні ніг і може пробігати декілька метрів за раз по поверхні води[54].
Плавання |

Ропуха звичайна (Bufo bufo) пливе
Безхвості, що мешкають у воді, адаптовані до плавання; для будови їх тіла характерні задні кінцівки і наявність плавальних перетинок між пальцями. Перетинки збільшують площу поверхні ступні (подібно ластам) і сприяють швидкому пересуванню жаб у воді. Представники родини Pipidae ведуть повністю водний спосіб життя і тому краще за всіх пристосовані до цього середовища проживання. Вони мають негнучкий хребет, тулуб плескатий і витягнутий, міцні задні кінцівки мають великі перетинки, для кращої орієнтації у воді наявний орган бічної лінії[64]. Пуголовки, як правило, мають великі хвостові плавці, які надають прискорення, направлене вперед, коли хвіст рухається з боку у бік. У воді безхвості найбільш безпорадні під час метаморфозу, коли хвіст вже атрофований, а ноги ще не повністю функціональні[59].
Риття нір |

Пурпурова жаба
Деякі безхвості адаптувалися до життя під землею і, як наслідок, до риття нір. У цих видів безхвостих, як правило, округлий тулуб, короткі кінцівки, маленька голова з виряченими очима, а задні ноги адаптовані до риття. Прикладом високої спеціалізації в цьому напрямі є пурпурова жаба, південноіндійських вид. Вона харчується термітами і проводить під землею майже все своє життя. Вилазить вона на короткий період під час мусонів, коли в калюжах відбувається спарювання і розмноження. У цієї жаби крихітна голова з витягнутою мордою і округле тіло. Через свій підземний спосіб життя цей вид був вперше описаний лише в 2003 році, хоча місцевим жителям вона була знайома вже давно[65]
Представники родини Scaphiopodidae, що мешкають в Північній Америці, теж адаптовані до підземного життя. Типовий вид, жаба Spea bombifrons, має ороговілу ділянку на плеснах задніх ніг, яку вона використовує, щоб зариватися в землю. Копаючи, жаба крутить стегнами з боку в бік, занурюючись в пухку землю. Влітку вона задовольняється маленькою норою, з якої по ночах виходить на полювання, але взимку ця жаба заривається набагато глибше (до 4,5 метрів вглиб)[66]. Утворений тунель заповнює земля, а жаба впадає в зимову сплячку в дальньому кінці нори. Під час сплячки жаба отримує воду з навколишнього середовища шляхом осмосу, але, не маючи можливості позбуватися сечовини, накопичує її в організмі[66]. Навесні Scaphiopodidae залишають свої зимові нори практично одночасно і намагаються потрапити до калюж, де вони спарюються. Відповідне місце голосно «анонсується» першим самцем, що його знайшов[67].
Інший вид, австралійський Heleioporus albopunctatus, веде зовсім інший спосіб життя. Ця жаба риє нору на березі чи під дном ріки і регулярно вилазить за їжею. Парування і відкладення ікри відбувається в гнізді всередині нори. Ікринки розвиваються до певної стадії, але пуголовки не залишають їх допоки нору не заллє сильною зливою. Тільки тоді пуголовки випливають у відкриту воду, де швидко завершують свій розвиток[68]. Мадагаскарські жаби з роду Scaphiophryne зариваються у сухе листя. Один з представників цього роду, Scaphiophryne marmorata, має сплюснуту голову і добре розвинені плеснові нарости на задніх ногах, що допомагають йому закопуватися. На передніх ногах цієї жаби є збільшені пальцеві диски, які допомагають їй пробиратися крізь кущі[69]. Цей вид жаб розмножується у калюжах, що з'являються після дощів[70].
Лазіння по деревах |

Phyllomedusa burmeisteri
Деревні безхвості мешкають у кронах дерев, де вони лазять по гілках, голках і листю. Деякі з них взагалі ніколи не спускаються на землю. «Справжні» деревні жаби відносяться до родини Hylidae, але зустрічаються представники інших родин безхвостих, адаптовані до деревного способу життя.

Ілюстрація жаби Rhacophorus nigropalmatus, що планерує, з книги Альфреда Воллеса «Малайський архіпелаг, країна орангутанга і райського птаха».
Так, серед дереволазів зустрічаються представники родин скляних жаб, Hyperoliidae, Microhylidae і Rhacophoridae[59]. Більшість деревних безхвостих не більше 10 см у довжину і можуть упіймати комаху, повиснувши на гілці вниз головою на одному пальці, або сидячі на тростинці, що гойдається під вітром[71]. У деяких представників підродини Phyllomedusinae на ногах є проставлені пальці. Це дозволяє їм хапатися за стеблини прибережних рослин[72].
Планерування |
Протягом своєї еволюційної історії декілька не пов'язаних між собою видів безхвостих адаптувались до планерування[73]. Деякі види жаб у тропічних лісах спеціалізовані для планерування з дерева на дерево або мають можливість контрольовано стрибати з дерева на землю («парашутування»). Типовими представниками таких безхвостих є Rhacophorus nigropalmat, що мешкає в Малайзії і Борнео. У цього виду великі ступні, кінчики пальців розширені і мають клейкі подушечки, між пальців ніг є літальні перетинки, а вздовж кінцівок в області таза наявні додаткові складки шкіри. Розтягуючи пальці і кінцівки, ця жаба може планерувати на значні відстані (до 15 метрів) між дерев, змінюючи у міру необхідності напрям руху[74][75].
Самозахист |

Не отруйна жаба Ranitomeya imitator
На перший погляд, безхвості виглядають досить беззахисними у світлі їх невеликих розмірів, повільного пересування, тонкої шкіри і відсутності захисного приладдя (наприклад, рогів, зубів і кігтів). Багато безхвостих мають забарвлення нейтральних кольорів, що дозволяє бути непомітними на тлі довкілля (поки жаба нерухома). Інші здатні робити великі стрибки з суходолу у воду, що дозволяє їм втікати від хижаків[76].
Багато безхвостих продукують токсичні речовини (буфотоксини), які роблять їх неїстівними для неспеціалізованих хижаків. Більшість ропух і деякі жаби мають великі паротоїдні залози, розташовані позаду очей, що виділяються слиз і токсини, які роблять безхвостих одночасно слизькими і отруйними, Якщо ефект отруєння відчувається моментально, хижак може випустити жабу. Якщо отрута має уповільнену дію, впійману жабу вона не врятує, але хижак (якщо вживе) у подальшому уникатиме представників даного виду[77].

Ропуха звичайна в захисній позі
Отруйні безхвості, як правило, сигналізують про свою токсичність за допомогою яскравого забарвлення шкіри (адаптивна стратегія, під назвою апосематизм). Деякі неотруйні види мімікрують під отруйних. Так, наприклад, жаба Allobates zaparo неотруйна, але мімікрує під два різних види, які мешкають на її території. У випадку присутності обох видів разом, Allobates zaparo копіює менш токсичну[78].
Деякі види поєднують маскувальну стратегію з апосематизмом. Так, кумка червоночерева має нейтральне забарвлення верхньої частини тіла і яскраві червоні чи жовті плями на череві. У разі небезпеки кума прибирає позу, що демонструє яскраві плями, і відлякує хижака[79].
Деякі безхвості у таких ситуаціях симулюють загрозу. Ропуха звичайна прибирає характерну стійку — роздувається, опускає голову і задирає зад[80]. Жаба-бик у небезпеці присідає, прикриває очі і схиляє голову вперед. Ця поза випинає партоїдні залози і закриває вразливі частини її тіла[76]. Деякі жаби у момент загрози видають гучний проникливий звук, який може відлякати хижака[76]. Незважаючи на те, що більшість хижаків уникає жаб, останні входять до раціону підв'язкових змій. Спостереження за поведінкою Anaxyrus americanus, які нещодавно пройшли метаморфоз, свідчать, що побачивши змію вони присідають і завмирають. Як правило, ця стратегія себе виправдовує — змія проповзає повз і не помічає жабу[81].
Життєвий цикл |
Розмноження |

Розвиток зародків Rana temporaria в ікринках протягом 11 днів.
У безхвостих спостерігаються два тими шлюбної поведінки: сезонна і спонтанна. При першому типі, характерному для більшості видів безхвостих, в певні періоди року дорослі жаби збираються для розмноження в ставках, озерах або струмках. Багато безхвостих повертаються до тієї водойми, в якій вони розвивалися пуголовками. У результаті можна спостерігати сезонні міграції, в які залучені тисячі особин. При спонтанному типі розмноження дорослі жаби прибувають у місця розмноження під поштовхом якоїсь зовнішньої причини (наприклад, випадінням опадів в посушливій місцевості). В такому випадку розмноження і розвиток ікринок та пуголовків відбувається з максимальною швидкістю, до того, як недовговічні водойми (калюжі) встигнуть висохнути[76].
Самці видів безхвостих з сезонним типом шлюбної поведінки звичайно першими прибувають до місць розмноження і залишаються там тривалий час, вони закликають кваканням самиць і захищають свою територію від інших самців. Самиці з'являються пізніше лише на час парування і викидання ікри. В цій ситуації кількість самців біля поди завжди перевищує кількість самиць, тому серед самців існує жорстка конкуренція. Великі і сильні самці видають більш низькі звуки і захоплюють найпривабливіші для розмноження місця. Спостереження показали, що самиці віддають перевагу саме таким самцям[82]. У деяких видів самці можуть застосовувати техніку перехоплення. У них немає своєї території і вони не квакають, а перехоплюють самиць, що знаходять на шляху до самця, що кличе самиць. Оскільки квакання потребує великих затрат енергії, іноді ролі змінюються і самець, що кликав, залишає свою територію і стає перехоплювачем[76].
у видав зі спонтанним типом шлюбної поведінки (наприклад, роду лопатоногів Scaphiopus) основний упор робиться не на вибір партнер, а на швидкість розмноження. Відповідно змінюється й тактика. Самець, що першим знаходить придатне місце (калюжу) починає гучно кликати родичів, на його заклик збігаються всі навколишні представники обох статей, які одразу підхоплюють заклик, створюючи чутний здалеку хор. Іноді придатні умови відсутні роками, і, відповідно, ці види не розмножуються декілька років поспіль[76]. Самиці виду Spea multiplicata звичайно відкладають за один шлюбний сезон близько половини наявної в них ікри. Приускають, що ця стратегія розрахована на появу більш слушних умов для розмноження у майбутньому[83].

Самець і самиця ропухи звичайної в амплексусі
За винятком деяких представників роду дереволазів, що утворюють на час шлюбного сезону моногамні пари, всі безхвості полігамні[84].
Як правило, процес парування полягає у тому, що самець сідлає самицю у воді і обхоплює її тіло. Самиця відкладає ікру, а самець покриває її своєю спермою (зовнішнє запліднення). У деяких видів самці можуть затримувати ікринки задніми лапами на час, необхідний для запліднення[76]. у цьому плані унікальним серед безхвостих є види західноафриканського роду Nimbaphrynoides, які розмножуються живородінням, і види танзанійського роду Nectophrynoides, що практикують яйцеживородіння. В обох випадках має місце внутрішнє запліднення, а самиця народжує цілком розвинутих молодих жабенят (сеголеток), які обминають стадію пуголовка[85][86].
Ікра |

Жаб'яча ікра
Жаб'яча ікра, як правило, запаковано у багатошаровий желатиновий матеріал, що забезпечує ікринкам певний захист і не перешкоджає проходженню (кисню, вуглекислого газі і аміаку). Ця захисна оболонка поглинає вологу і розбухає у воді. Після запліднення внутрішня частина ікринки розріджується, що забезпечує зародку свободу руху при розвитку. У деяких видів (наприклад, червоноокої жаби і Rana sylvatica) у желатиновому матеріалі наявні одноклітинні зелені водорості. Було висунуте припущення, що вони справлять позитивний вплив на розвиток зародка, підвищуючи концентрацію кисню, який утворюється у процесі фотосинтезу[87]. У більшості видів ікринки мають чорний або темно-коричневий колір, що дозволяє їм нагріватись під променями сонця сильніше навколишнього середовища. Так, наприклад, температура всередині кластера ікри Rana sylvatica була на 6 °C вище температури води, що сприяло швидкому розвитку зародків[88]
Розмір і форма аґломерату ікри характерна для кожного виду. Жаби родини Ranidae мають тенденцію до кулястих кластерів, а ропухові до довгих циліндричних кластерів. Крихітний кубинський свистун відкладає по одній ікринці і закопує їх у вологий грант[89]. Leptodactylus pentadactylus створює пінне гніздо у норі, відкладаючи у нього близько тисячі ікринок. Пуголовки з'являють на світ, коли вода заповнює нору, а іноді розвиток повністю проходить у гнізді[90]. Agalychnis callidryas з родини райкові відкладає ікринки на листя, розташоване над поверхнею водойми. Після вилуплення пуголовки падають з листя у воду[91].
У деяких видів на певній стадії розвитку зародки у ікринках можуть вловити вібрації, що спричинюються хижаками (осами, зміями), і вилупитися раніше часу, щоб дістати мобільність і уникнути гибелі[92]. Загалом, тривалість стадії розвитку зародків в ікрі залежить від конкретного виду і навколишніх умов. Як правило, пуголовки вилуплюються протягом тижня після того, як капсула ікринки розпадається під дією гормону, що виробляється зародком[93].
Пуголовки |

Життєвий цикл Rana clamitans
Личинки безхвостих, що з'являються з ікринок, відомі як «пуголовки». Вони ведуть повністю водний спосіб життя, але відоме один виняток — пуголовки виду Nannophrys ceylonensis напівсухопутні і мешкають серед мокрого каміння[94][95]. Тіло пуголовків, як правило, має овальну форму, хвіст довгий, сплющений по вертикалі, пристосований для плавання. У пуголовків хрящовий хребет, очі не мають повік, є орган бічної лінії, для дихання слугують зябра[96].
Спочатку у пуголовків з'являються зовнішні зябра, а пізніше внутрішні (зябровий мішок закриває зябра і передні ноги). Легені, що розвиваються, слугують додатковим дихальним органом. Деякі види проходять метаморфоз ще в ікринці і з'являються у світ вже жабенятами. У пуголовків немає справжніх зубів, але у багатьох видів щелепи вкриті паралельними рядами кератинових зубчиків (два ряди на верхній щелепі, три ряди на нижній і роговий дзьоб). Кількість рядів і точна морфологія варіюють у різних видів і можуть слугувати діагностичною ознакою[93]. Пуголовки родини пипові (за винятком роду Hymenochirus) мають перу передніх вусиків, що роблять їх схожими на невеликих сомиків[64].
Пуголовки, як правило травоїдні і харчуються водоростями, відфільтрованими з води через зябра. Деякі види є хижаками вже на стадії пуголовка і харчуються комахами; пуголовки (Osteopilus septentrionalis практикують канібалізм, а також поїдають маленьких риб. Пуголовки, що рано відростили ноги, можуть стати жертвою своїх співбратів[97].
На пуголовків полюють риби, саламандри, хижі жуки і птахи (наприклад, рибалочка). Деякі пуголовки, у тому числа личинки Bufo marinus, отруйні. У різних видів безхвоста стадія пуголовка триває від тижня до декількох місяців і залежить від стратегії розмноження[98].
Метаморфоз |
Після завершення стадії пуголовка безхвості проходять процес метаморфозу, під час якого системи організму різко перебудовують у дорослу форму. Як правило метаморфоз триваю близько доби. Він починається з вироблення гормону тироксину, який впливає на розвиток тканин. Так, зміна органів дихання включає розвиток легенів паралельно зі зникненням забрів і зябрового мішку. Передні кінцівки стають видимими. Нижня щелепа набуває характерної для хижака форми, кишечник стає коротшим. Нервова система адаптується під стереоскопічні зір і слух, а також під нові способи пересування і харчування. Очі зсуваються вище, формуються повіки і пов'язані з ними залози. Утворюються органи слуху (слухова перетинка, середнє і внутрішнє вухо). Шкіра стає більш товстою і міцнішою, орган бічної лінії зникає (у більшості видів), формують шкіряні залози. На останньому етапі метаморфозу зникає хвіст, його тканини йдуть на розвиток кінцівок[99][100].

Личинка жаби Rana temporaria за день до метаморфозу.

Середина метаморфозу — щелепи трансформовані, очі збільшені, видно залишки зябрового мішку.

Жабеня із залишками хвосту, метаморфоз майже завершений.



Дорослі особини |
Безхвості, що пройшли метаморфоз, переходять у характерне для свого виду середовище проживання. Практично всі види дорослих безхвостих — хижаки. Вони полюються на безхребетних, включно з членстоногими, червами і равликами. Існують великі види поїдають інших земноводних, невеликих ссавців і птахів. Одні безхвості ловлять свою жваву здобич клейким язиком, інші запихають їжу до рота передніми кінцівками. Деревна жаба Xenohyla truncata є винятком, оскільки харчується фруктами[101]. На безхвостих полють багато хижаків, у тому числі чаплі, яструби, риби, великі саламандри, змії, єноти, тхори та інші[102].
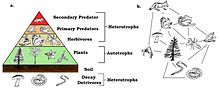
Екологічна піраміда. Безхвості показані як первинні хижаки.
Безхвості є первинними хижаками, важливою складовою харчового ланцюжку. Ці холоднокровні тварини ефективно споживають їжу — витрачають лише невелику частину енергії на метаболічні процеси, а решту конвертують в біомасу. Вони слугують їжею вторинним хижакам, а самі харчуються членистоногими, здебільшого травоїдними. Таким чином, поїдаючи споживачів рослин, безхвості збільшують приріст біомаси рослин, що сприяє балансу екосистеми[103].
Тривалість життя безхвостих у природних умовах є маловивченою. З використанням методів хребтохронології тривалість життя жаби Rana muscosa була виміряна за сезонними змінами росту фаланг пальців. Отримані дані дозволяють зробити припущення, що максимальна тривалість життя дорослої особини становить 10 років, а враховуючи стадію пуголовка, яка у цього виду триває близько 4 років, тривалість життя цих жаб рівний 14 рокам[104]. У неволі зареєстрована тривалість життя безхвостих до 40 років, такого віку досягла ропуха звичайна. Жаба-ага жила у неволі 24 роки, а жаба-бик — 14 років[105].
Піклування про потомство |

Маленький древолаз

Жаба-повитуха (самець) з ікрою

Сумчаста жаба
Способи піклування про потомство у безхвостих вивчені недостатньо. Припускають, що близько 20% видів земноводних так чи інакше піклуються про дитинчат[106]. Існує зворотний зв'язок між розмірами водойми, що слугує для розмноження, і рівнем батьківської опіки, яку проявляють безхвості. Види безхвостих, що розмножуються в невеликих водоймах, демонструють більш складні рівні батьківського піклування[107]. У великих водоймах великий процент ікри і пуголовків поїдається хижаками. Та цьому тлі деякі види безхвостих адаптувались відкладати ікру на суходолі. Зокрема вони піклуються про те, щоб підтримувати ікру вологою в умовах суходолу[108]. Додаткова батьківська опіка проявляється при транспортуванні до водойм пуголовків, що вилупились на суходолі[107].
У невеликих водоймах хижаків менше і виживання пуголовків регулюється в основному внутрішньою конкуренцією. Деякі види жаб для уникнення цієї конкуренції переносять нових пуголовків у невеликі внутршньорослинні порожнини (лат. phytotelmata), заповнені водою[109]. Незважаючи на відсутність конкуренції, в таких порожнинах мало ресурсів, тому батькам доводиться годувати своїх пуголовків. Деякі види годують пуголовків незаплідненими клітинами[107]. Приміром, маленький дереволаз (Oophaga pumilio) відкладає ікру просто на землі у лісі. Самець охороняє ікру від хижаків і зволожує її з клоаки, щоб вона не пересохла. Коли пуголовки вилуплюються, самиця переносить їх на сині у порожнину якого-небудь представника родини бромелієвих, залишаючи у кожній рослині по одному пуголовку. Після цього самиця регулярно відвідує пуголовків, відкладаючи кожному по одній-двом незаплідненим ікринкам для споживання як їжі, і продовжує їх годувати до настання метаморфозу[110]. Подібним способом піклуються про своє потомство й представники виду Oophaga granulifera[111].
Форми батьківського піклування у безхвостих вельми різноманітні. Крихітний самець Colostethus subpunctatus охороняє свій кластер ікри, відкладений під каменем чи деревиною. Коли пуголовки вилуплюються, він переносить їх на спині (приклеєними слизовими виділеннями) у тимчасову водойму, де, частково занурюючись у воду, звільняє одного чи декількох пуголовків, після чого переходить на наступної водойми[112]. Самець жаби-повитухи носить ікринки з собою приклеєними до задніх кінцівок. У посушливу погоду він зволожує їх, занурюючись у ставок, а щоб вони занадто не намокнули у мокрій траві, підіймає задню частину тіла. Через три-шість тижнів він перестає виходити зі ставка, і за деякий час після цього з ікринок вилуплюються пуголовки[113]. Південноамериканська жаба Engystomops pustulosus споруджує з піни гніздо, в яке відкладає ікру. Піноматеріал складається з білків і лектинів і, можливо, має антибактеріальні властивості[114]. Декілька пар жаб можуть будувати спільне гніздо. У цьому випадку спершу будується «пліт», а потім жаби відкладають ікру в його центрі, при цьому чергуються відкладання ікри і формування піноматеріалу, а завершують процес створенням шару піни зверху ікри[115].
Деякі види безхвостих зберігають потомство всередині своїх тіл. Самець і самиця сумчастої жаби разом охороняють ікру, відкладену на землі. Коли настає час вилуплення пуголовків, самець змащує своє тіло слизовим матеріалом, що оточує ікринки, і занурюється в їх масу. Після вилуплення пуголовки влізають у шкіряні кишені самця, де залишаються до завершення метаморфозу[116]. Самиці реобатрахусів (можливо нещодавно вимерлі) проковтували свої запліднені ікринки, які розвивались у них в шлунку. На цей час жаби припиняли їсти і виділяти шлункові соки, а пуголовки харчувались жовтком з ікринок. Через шість-сім тижнів самиці широко розкривали рот і відригали пуголовків назовні[117].
Самиця виду рінодерма Дарвіна, що мешкає в Чилі, відкладає на землю 40 ікринок, які охороняють самцем. Коло пуголовки вилуплюються, самець заковтує їх і тримає їх у своєму збільшеному горловому мішку. Пуголовки занурені у в'язку пінисту рідину, яка додатково до жовтка постачає їх поживні речовини. Вони залишаться у мішку від семи до десяти тижнів, після чого проходять метаморфоз, перемірюються у ротову порожнину самця і виплигують назовні[118].
Використання людиною |
Утримання в неволі |
Як домашніх тварин любителі земноводних часто утримують безхвостих у тераріумах, акватераріумах і акваріумах[119]. Безхвостих також розводять і використовують як живий корм для деяких мешканців «домашніх зоопарків» або «живих кутків».
В кулінарії |

Приготовлене жаб'яче м'ясо. Цзяньян, Фуцзянь, Китай
Жаб'ячі лапки в різних країнах уживають як їжу. Традиційний спосіб задоволення потреб місцевого ринку за рахунок локальних популяцій останніми рокам став неможливим через скорочення цих популяцій. В теперішній час існує розвинена міжнародна торгівля жаб'ячими лапками. Основні імпортери — Франція, Бельгія, Люксембург і США, основні експортери — Індонезія і Китай[120]. Річний обіг продажів американської жаби-бика (Rana catesbeiana), яку промислово розводять у Китаї, досягає 2,4 тисяч тон[121]. Розведення їстівних жаб останнім часом цікавляться й інші країни, наприклад, Білорусь[122].
В науково-дослідній сфері |
Безхвостих широко використовували в наукових експериментах. У XVIII столітті біолог Луїджі Гальвані відкрив за допомогою експериментів з жабами зв'язок між електрикою і нервовою системою[123]. У 1852 році Г. Ф. Станніус використовував серце жаби в експерименті, названому його ім'ям, який довів, що шлуночки серця і передсердя мають незалежний один від одно ритм[124]. шпоркову жабу гладеньку широко використовували в першій половині XX століття в тесті на вагітність після того, як англійський зоолог Ланселот Хогбен відкрив, що в сечі вагітної жінки присутній гормон хоріонічний гонадотропін, який індукує цю жабу викидати ікру[125]. У 1952 році Роберт Бріггс і Джозеф Кінг клонували жабу методом пересадки ядер соматичних клітин (цим же методом пізніше була клонована вівця Доллі). Це був перший успішний експеримент з клонування вищих тварин шляхом ядерної трансплантації[126]. Жаб широко використовують в дослідженнях в галузі ембріології. Шпоркові жаби залишились модельними організмами в біології розвитку й після розроблення сучасніших тестів на вагітність, оскільки їх легко утримувати в лабораторних умовах, а їх ембріони достатньо великі для маніпулювання[127]. При цьому шпоркових жаб гладеньких частіше замінюють дрібнішим родичем, Xenopus tropicalis, який досягає статевої зрілості у 5 місяців (а не в рік або два, як шпоркова жаба гладенька)[128], і це прискорює дослідження, що потребують декількох поколінь жаб. Геном X. tropicalis станом на 2012 рік перебував у процесі секвенування[129].
Надзвичайна різноманітність токсинів, які виробляються безхвостими, викликала інтерес біохіміків до цієї «природної аптеки». Алкалоїд епібатидин, болезаспокійливе, у 200 разів більш потентне, ніж морфій, був виявлений у деяких видів роду листолазів. Зі шкіри безхвостих був виділений пептид, що можливо блокує ділення вірусу ВІЛ[130][131].
Жаб використовують на практичних заняттях з диссекції в школах та університетах. Як правило, їх попередньо обробляють пігментами для отримання контрасту між різними системами організму. На тлі руху за права тварин цю практику останнім часом заступає віртуальна диссекція оцифрованих жаб — комп'ютерні програми, що симулюють організм живої жаби[132].
Виробництво отрути |
Отруту безхвостих застосовують як галюциноген. Тривалий час вважалось, що у доколумбову добу жителі Мезоамерики використовували з цією метою отруту жаби-аги. Пізніші дослідження показали, що більш вірогідним кандидатом на цю роль є отрута жаби Bufo alvarius, що містить буфотенін (5-MeO-DMT), психоактивну речовину, яку й нині застосовують у рекреаційних цілях. Типовим способом вживання є куріння висушених шкіряних виділень жаби[133]. У ЗМІ з'являлись повідомлення про людей, котрі злизували наркотичну речовину просто зі шкіри жаби, але, можливо, це міська легенда[134].

Жахливий листолаз (Phyllobates terribilis)
Крім того, отруту безхвостих здавна застосовували для виготовлення отруйних стріл і дротиків. За допомогою шкіряних виділень жахливого листолаза південноамериканські індійці виготовляли отруйні стрілки. Наконечник натирали об спину жаби, стрілками стріляли з духової трубки на полюванні. Комбінація двох токсинів, наявних y цих виділеннях (батрахотоксин і гомобатрахотоксин), є настільки сильною, що отрути однієї жаби, як гадають, достатньо, щоб вбити 22 000 мишей[135]. Два інших види листолазів (Phyllobates aurotaenia і Phyllobates bicolor), також використовували як джерело отрути, але концентрація отрути в них менше, і для того, щоб вона почала виділятись, їх потрібно підігріти на вогні[136]. Ці отрути досліджують щодо можливості використання в медицині[137].
Охоронний статус |
Дослідження, розпочаті у 1950-х роках, відмічають значне скорочення чисельності безхвостих. Більше третини видів перебувають під загрозою вимирання, понад 120 видів вважаються вимерлими в період з 1980 по 2004 рік[138]. Серед цих видів значиться й помаранчева жаба з Коста-Рики. Її зникнення викликало серйозне занепокоєння дослідників, пов'язане з тим, що ця жаба мешкала у заповіднику «Monteverde Cloud Forest Reserve» і її вимирання не могло бути викликане безпосередньо людським втручанням. В інших місцях зниження чисельності безхвостих зумовлене руйнуванням середовища проживання, полютантами, змінами клімату і поширенням чужорідних хижаків, паразитів і конкурентів. Особливо руйнівним для популяції безхвостих вважаються інфекційні хвороби хітридіомікоз і ранавірус[2][139][140].
Багато дослідників вважають, що підвищена чутливість земноводних і, зокрема, безхвостих, до забруднення довкілля пов'язана з такими факторами, як їх проміжна позиція в харчовому ланцюжку, проникна шкіра і життєвий цикл, що включає водну стадію (пуголовок) і наземний спосіб життя дорослої особини[141]. Ті нечисленні види безхвостих, у яких водний етап життя скорочений або зовсім відсутній, мають більшу опірність забрудненню ніж типові безхвості, що розвиваються у воді від стадії ікринки до кінця метаморфозу[142].
Кількість мутацій і генетичних дефектів, що спостерігаються у безхвостих, збільшилась за час спостережень з 1990-х по 2003 рік. Одним з розповсюджених дефектів є відсутні або зайві кінцівки. Серед різноманітних гіпотез з питання причин, що викликають ці дефекти, — збільшення ультрафіолетового випромінювання, що потрапляє на ікру, забруднення сільськогосподарськими отрутохімікатами і паразитичні захворювання, наприклад, зараження трематодами Ribeiroia ondatrae. Можливо, що всі ці фактори діють разом (радіаційний і хімічний стрес знижують опірність організмів паразитам). Дефекти кінцівок погіршують рухливість і, відповідно, шанси тварини дожити до статевої зрілості[143][144].
Дослідження, проведене в Канаді у 2006 році, показало, що висока щільність траспорту для безхвостих є більшою загрозою, ніж погіршення середовища проживання[145]. В деяких випадках були започатковані програми розмноження в неволі, які, здебільшого були успішними[146][147]. У 2007 році було опубліковане дослідження, яке виявило, що деякі пробіотичні бактерії можуть підвищувати опірність безхвостих летальним грибковим захворюванням[148]. Була розроблена програма «Панамський проект зі спасіння і збереження амфібій» (англ. The Panama Amphibian Rescue and Conservation Project) для збереження у східній Панамі деяких видів безхвостих, які гинуть від цих хвороб; програма включає розробку польових методів використання пробіотиків[149]. Світова асоціація зоопарків і акваріумів (англ. World Association of Zoos and Aquariums) оголосила 2008 рік роком жаби з метою привернення уваги суспільства до проблеми охорони безхвостих[150].
Безхвості в культурі |

Скульптура жаби, Київ
У фольклорі |
У багатьох народів світу жаби, і ще у більшому ступені ропухи, асоціювались з цілою низкою неприємних властивостей[151]. В китайській традиції жаба символізує «Їнь». Дух жаби Цін-ва Шен асоціюється з одужанням і удачею у справах. Символ «жаба в колодязі» відноситься до людини обмеженої[152]. У європейському фольклорі ропухи асоціювались з відьмами як фамільяри, джерела інгредієнтів для магічного зілля, а іноді мали власні магічні властивості[153]. В давньоперуанській культурі Моче безхвості були одним з тварин, яким віддавали шану, а також часто з'являлись у творах мистецтва[154]. За легендою Панами, удача супроводжує того, хто побачить панамську золоту жабу (Atelopus zeteki). У варіанті цієї легенди говориться, що, після смерті ці жаби перетворюються на золотий талісман вака[155].
В українській етнокультурі жаби фігурують в замовляннях і зашіптуваннях, відоме їх використання у народній медицині. З поведінкою жаб пов'язана низка народних прикмет, наприклад «багато жаб на стежках — на дощ», «жаби квакають — на дощ». У народній мові жаби часто виступали об'єктом негативної характеристики (наприклад, «Одступись, зелена жабо!»), а також прокльону. Іронізування й кепкування спричиняють відсутність у жаби хвоста («якби жабі хвоста, була б непроста»), великий рот («жаба і на вола рот роззявляє»), а також характерне надування і квакання («жаба дулася, дулася, та й лопнула»). З цими тваринами пов'язано багато народних прислів'їв (наприклад, «кожна жаба своє болото хвалить»), приказок («роздайся, море, жаба лізе»)[156].
В літературі |
Безхвості часто виступають як персонажі в літературі[157]. Найдавніший художній твір, в назві якого фігурують безхвості і що дійшов до нашого часу, — це комедія Аристофана «Жаби», вперше поставлена у 405 році до н. е. Як додаткові приклади можна навести:
Батрахоміомахія — написана гекзаметром давньогрецька пародійна поема про війну мишей і жаб.- Царівна-жаба, жінка-жаба — персонажі слов'янських народних казок[158].
- Король-жабеня — персонаж казки братів Грімм «Казка про короля-жабеня, або про Залізного Генріха»[159].
Примітки |
↑ David C. Blackburn, David B. Wake. Class Amphibia Gray, 1825 // Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness / Zhi-Qiang Zhang (editor). — Auckland : Magnolia Press, 2011. — P. 39—55. — (Zootaxa) — ISBN 1175-5334.
↑ аб Маслова И. В. Влияние климата на отдельные аспекты жизнедеятельности земноводных и пресмыкающихся / Сост. А. О.Кокорин. — Москва : WWF России, 2006. — С. 111. — ISBN 5895640370. — Bibcode:.
↑ аб Marjanović D.,Laurin M. (2007). Fossils, Molecules, Divergence Times, and the Origin of Lissamphibians. Syst Biol 56(3: 369–388. doi:10.1080/10635150701397635.
↑ Alexander Pyron, R. (2011). Divergence Time Estimation Using Fossils as Terminal Taxa and the Origins of Lissamphibia. Syst Biol. 60(4): 466–481. doi:10.1093/sysbio/syr047.
↑
Weishampel, D. B.; Dodson, P.; Osmólska, H. (eds.) (2004). Dinosaur distribution (Early Jurassic, North America): The Dinosauria (2nd edition). University of California Press. с. 530‑532. ISBN 9780520242098.
↑ Grzimek, 2003, с. 11
↑ Shubin, N. H.; Jenkins, F. A. Jr (1995). An Early Jurassic jumping frog. Nature 377 (6544): 49‑52. doi:10.1038/377049a0.
↑ Foster, J. (2007). Anura (Frogs). Jurassic west: the dinosaurs of the Morrison Formation and their world. Indiana University Press. с. 135‑136. ISBN 0-253-34870-6.
↑ David Cannatella, David M. Hillis. Amphibians: Leading a Life of Slime // Assembling the Tree of Life / Edited by Joel Cracraft, Michael J. Donoghue. — Oxford University Press, 2004. — P. 438. — ISBN 0-19-517234-5.
↑ Evans, S. E.; Jones, M. E. H.; Krause, D. W. (2008). A giant frog with South American affinities from the Late Cretaceous of Madagascar. Proceedings of the National Academy of Sciences 105 (8): 2951‑2956. PMC 2268566. PMID 18287076. doi:10.1073/pnas.0707599105.
↑ Freaky Frogs. National Geographic Explorer. Архів оригіналу за 2012-10-17. Процитовано 2012-07-13.
↑ Ferrell, Vance (2012-03-04). Geographical Distribution. Evolutio]12-10-17. Архів оригіналу за 2013-06-25.
↑ Introduced frogs. New Zealand Frog Research Group. 2006. Архів оригіналу за 2012-10-17. Процитовано 2012-10-07.
↑ Ryan, Paddy (2011-09-25). Story: Frogs. The Encyclopedia of New Zealand. Архів оригіналу за 2012-10-17. Процитовано 2012-08-20.
↑ Dahl, Chris; Novotny, Vojtech; Moravec, Jiri; Richards, Stephen J. (2009). Beta diversity of frogs in the forests of New Guinea, Amazonia and Europe: contrasting tropical and temperate communities. Journal of Biogeography 36 (5): 896‑904. doi:10.1111/j.1365-2699.2008.02042.x.
↑ Cyclorana platycephala. Frogs Australia Network. 2005-02-23. Процитовано 2012-07-20.
↑ абвгд Tesler, P. (1999). The amazing adaptable frog. Exploratorium:: The museum of science, art and human perception. Архів оригіналу за 2012-08-06. Процитовано 2012-06-04.
↑ Grzimek, 2003, с. 26
↑ аб Grzimek, 2003, с. 15
↑ абв Карташев Н.Н., Соколов В.Е., Шилов И.А. Практикум по зоологии позвоночных. zoomet.ru. Архів оригіналу за 2012-10-17. Процитовано 2012-10-07.
↑ Harvey, M. B, A. J. Pemberton, and E. N. Smith (2002). «New and poorly known parachuting frogs (Rhacophoridae : Rhacophorus) from Sumatra and Java». Herpetological Monographs 16: 46—92.
↑ Walker, M. (2009-06-25). Legless frogs mystery solved. BBC News.
↑ аб Badger, D.; Netherton, J. (1995). Frogs. Airlife Publishing. с. 27. ISBN 1853107409.
↑ аб Wente, W. H.; Phillips, J. B. (2003). Fixed green and brown color morphs and a novel color‐changing morph of the Pacific tree frog Hyla regilla. The American Naturalist 162 (4): 461‑473. JSTOR 10.1086/378253.
↑ Smyth, H. R. (1962). Amphibians and Their Ways. Macmillan. ISBN 0026121905.
↑ Grzimek, 2003, с. 83
↑ Grant, T. Dart Poison Frog Vivarium. American Museum of Natural History. Архів оригіналу за 2012-10-17. Процитовано 2012-10-14.
↑ Smith, B. P., Tyler M. J., Kaneko T., Garraffo H. M., Spande T. F., Daly J. W. (2002). «Evidence for biosynthesis of pseudophrynamine alkaloids by an Australian myobatrachid frog (pseudophryne) and for sequestration of dietary pumiliotoxins». J Nat Prod 65 (4): 439—47.
↑ Saporito, R.A., H.M. Garraffo, M.A. Donnelly, A.L. Edwards, J.T. Longino, and J.W. Daly (2004). «Formicine ants: An arthropod source for the pumiliotoxin alkaloids of dendrobatid poison frogs». Proceedings of the National Academy of Science 101: 8045—8050.
↑ Savage, J. M. (2002). The Amphibians and Reptiles of Costa Rica. University of Chicago Press, Chicago.
↑ Duellman, W. E. (1978). «The Biology of an Equatorial Herpetofauna in Amazonian Ecuador». University of Kansas Museum of Natural History Miscellaneous Publication 65: 1-352.
↑ Kimball, John (2010). Vertebrate Lungs: Frog Lungs. Kimball's Biology Pages. Архів оригіналу за 2012-08-06. Процитовано 2012-07-09.
↑ Boisvert, Adam (2007-10-23). Barbourula kalimantanensis. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-08-06. Процитовано 2012-07-09.
↑ Kimball, John (2010). Animal Circulatory Systems: Three Chambers: The Frog and Lizard. Kimball's Biology Pages. Архів оригіналу за 2012-08-06. Процитовано 2012-07-09.
↑ Lee, Deborah (2010-04-23). Telmatobius culeus. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-08-06. Процитовано 2012-07-09.
↑ абвг Frog's internal systems. TutorVista.com. 2010. Архів оригіналу за 2012-08-06. Процитовано 2012-06-04.
↑ Armstrong, Cecilia E.; Roberts, William M. (1998). Electrical properties of frog saccular hair cells: distortion by enzymatic dissociation. Journal of Neuroscience 18 (8): 2962—2973. PMID 9526013.
↑ Emmer, Rick (1997-11-24). How do frogs survive winter? Why don't they freeze to death?. Scientific American. Архів оригіналу за 2012-08-17. Процитовано 2012-06-15.
↑ Kayes, Sara M.; Cramp, Rebecca L.; Franklin, Craig E. (2009). Metabolic depression during aestivation in Cyclorana alboguttata. Comparative Biochemistry and Physiology - Part A: Molecular & Integrative Physiology 154 (4): 557‑563. doi:10.1016/j.cbpa.2009.09.001.
↑ Hudson, N. J.; Lehnert, S. A.; Ingham, A. B.; Symonds, B.; Franklin, C. E.; Harper, G. S. (2005). Lessons from an estivating frog: sparing muscle protein despite starvation and disuse. AJP: Regulatory, Integrative and Comparative Physiology 290 (3): R836‑R843. PMID 16239372. doi:10.1152/ajpregu.00380.2005.
↑ аб Cannatella, David (2008-01-11). Anura. Tree of Life web project. Архів оригіналу за 2012-10-17. Процитовано 2012-08-08.
↑ Ford, L.S.; Cannatella, D. C. (1993). The major clades of frogs. Herpetological Monographs 7: 94‑117. JSTOR 1466954. doi:10.2307/1466954.
↑ Faivovich, J.; Haddad, C. F. B.; Garcia, P. C. A.; Frost, D. R.; Campbell, J. A.; Wheeler, W. C. (2005). Systematic review of the frog family Hylidae, with special reference to Hylinae: Phylogenetic analysis and revision. Bulletin of the American Museum of Natural History 294: 1‑240. doi:10.1206/0003-0090(2005)294[0001:SROTFF]2.0.CO;2.
↑ Шабанов Д. А., Литвинчук С. Н. Зелёные лягушки: жизнь без правил или особый способ эволюции? // Природа. — 2010. — Вып. 3. — С. 29-36.
↑ Kuzmin, S. L. (1999-11-10). Rana esculenta. Архів оригіналу за 2012-10-17. Процитовано 2012-06-15.
↑ Bullfrog. Ohio Department of Natural Resources. Архів оригіналу за 2012-08-06. Процитовано 2012-06-19.
↑ Nafis, Gary (2012). Ascaphus truei: Coastal Tailed Frog. California Herps. Архів оригіналу за 2012-08-06. Процитовано 2012-08-01.
↑ Roy, Debjani (1997). Communication signals and sexual selection in amphibians (PDF). Current Science 72: 923‑927.
↑ Gerhardt, H. C. (1994). The evolution of vocalization in frogs and toads. Annual Review of Ecology and Systematics 25: 293‑324. doi:10.1146/annurev.es.25.110194.001453.
↑ абвг Badger, David; Netherton, John (1995). Frogs. Airlife Publishing Ltd. с. 39–44. ISBN 1853107409.
↑ Top 10 best jumper animals. Scienceray. Архів оригіналу за 2012-08-17. Процитовано 2012-06-11.
↑ James, R. S.; Wilson, R. S. (2008). Explosive jumping: extreme morphological and physiological specializations of Australian rocket frogs (Litoria nasuta). Physiological and Biochemical Zoology 81 (2): 176‑185. PMID 18190283.
↑ Nauwelaerts, S.; Schollier, J.; Aerts, P. (2004). A functional analysis of how frogs jump out of water. Biological Journal of the Linnean Society 83 (3): 413‑420. PMID 18190283. doi:10.1111/j.1095-8312.2004.00403.x.
↑ аб Badger, David; Netherton, John (1995). Frogs. Airlife Publishing. с. 51. ISBN 1853107409.
↑ Flam, F. (1995). Finding earliest true frog will help paleontologists understand how frog evolved its jumping ability. Knight Ridder/Tribune News Service via HighBeam Research. Архів оригіналу за 2012-08-17. Процитовано 2012-06-10.
↑ Minott, Kevin (2010-05-15). How frogs jump. National Geographic. Архів оригіналу за 2012-08-17. Процитовано 2012-06-10.
↑ Astley, H. C.; Roberts, T. J. (2011). Evidence for a vertebrate catapult: elastic energy storage in the plantaris tendon during frog jumping. Biology Letters 8 (3): 386–389. doi:10.1098/rsbl.2011.0982.
↑ Scott, J. (2005). The locust jump: an integrated laboratory investigation. Advances in Physiology Education 29 (1): 21‑26. doi:10.1152/advan.00037.2004.
↑ абв Anura. Encyclopædia Britannica Online. Архів оригіналу за 2012-08-17. Процитовано 2012-06-12.
↑ Fitch, H. S. (1956). An ecological study of the collared lizard (Crotaphytus collaris). University of Kansas Publications (Museum of Natural History) 8: 213‑274.
↑ Walton, M.; Anderson, B. D. (1988). The aerobic cost of saltatory locomotion in the fowler's toad (Bufo woodhousei fowleri). Journal of Experimental Biology 136: 273‑288.
↑ Ahn, A. N.; Furrow, E.; Biewener, A. A. (2004). Walking and running in the red-legged running frog, Kassina maculata. Journal of Experimental Biology 207: 399‑410. doi:10.1242/jeb.00761.
↑ Pickersgill, M.; Schiøtz, A.; Howell, K.; Minter, L. (2004). Kassina maculata. IUCN Red List of Threatened Species. Архів оригіналу за 2012-08-17. Процитовано 2012-06-11.
↑ аб Pipidae. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-08-17. Процитовано 2012-06-14.
↑ Radhakrishnan, C.; Gopi, K. C. (2007). Extension of range of distribution of Nasikabatrachus sahyadrensis Biju & Bossuyt ( Amphibia : Anura : Nasikabatrachidae ) along Western Ghats , with some insights into its bionomics. Current Science 92 (2): 213‑216. ISSN 00113891.
↑ аб Farrar, Eugenia; Hey, Jane. Spea bombifrons. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-08-17. Процитовано 2012-06-16.
↑ Scaphiopodidae. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-08-17. Процитовано 2012-06-16.
↑ Roberts, Dale; Hero, Jean-Marc (2011). Heleioporus albopunctatus. IUCN Red List of Threatened Species. Архів оригіналу за 2012-08-17. Процитовано 2012-06-16.
↑ Staniszewski, Marc (1998-09-30). Madagascan Burrowing Frogs: Genus: Scaphiophryne (Boulenger, 1882). Архів оригіналу за 2012-08-17. Процитовано 2012-06-16.
↑ Venesci, M; Raxworthy, C. J.; Nussbaum, R. A.; Glaw, F. (2003). A revision of the Scaphiophryne marmorata complex of marbled toads from Madagascar, including the description of a new species. Herpetological Journal 13: 69‑79.
↑ Cochran, Doris Mabel (1961). Living Amphibians of the World. Doubleday. с. 112. ISBN 9780241903384.
↑ Phyllomedusa ayeaye. AmphibiaWeb. Архів оригіналу за 2012-08-17. Процитовано 2012-06-14.
↑ Emerson, Sharon B.; Koehl, M. A. R. (1990). The interaction of behavioral and morphological change in the evolution of a novel locomotor type: "flying frogs. Evolution 44 (8): 1931‑1946. JSTOR 2409604.
↑ Shah, Sunny; Tiwari, Rachna (2001-11-29). Rhacophorus nigropalmatus. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-08-17. Процитовано 2012-06-11.
↑ Wallace's Flying Frog Rhacophorus nigropalmatus. National Geographic: Animals. Архів оригіналу за 2012-08-17. Процитовано 2012-06-05.
↑ абвгдеж Stebbins, Robert C.; Cohen, Nathan W. (1995). A Natural History of Amphibians. Princeton University Press. с. 154‑162. ISBN 0-691-03281-5.
↑ Barthalmus, George T.; Zielinski, William J. (1988). Xenopus skin mucus induces oral dyskinesias that promote escape from snakes. Pharmacology Biochemistry and Behavior 30 (4): 957‑959. doi:10.1016/0091-3057(88)90126-8.
↑ Darst, Catherine R.; Cummings, Molly E. (2006). Predator learning favours mimicry of a less-toxic model in poison frogs. Nature 440: 208‑211. doi:10.1038/nature04297.
↑ Kuzmin, Sergius L. (1999-09-29). Bombina bombina. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-10-17. Процитовано 2012-06-15.
↑ Arnold, Nicholas; Ovenden, Denys (2002). Reptiles and Amphibians of Britain and Europe. Harper Collins Publishers. с. 73‑74. ISBN 0002199645.
↑ Hayes, Floyd E. (1989). Antipredator behavior of recently metamorphosed toads (Bufo a. americanus) during encounters with garter snakes (Thamnophis s. sirtalis). Copeia (American Society of Ichthyologists and Herpetologists) 1989 (4): 1011‑1015. JSTOR 1445987.
↑ Davies, N. B.; Halliday, T. R. (1978). Deep croaks and fighting assessment in toads Bufo bufo. Nature 274: 683‑685. doi:10.1038/274683a0.
↑ Long, David R. (1989). Energetics and reproduction in female Scaphiopus multiplicatus from Western Texas. Journal of Herpetology 23 (2): 176‑179. JSTOR 1564026.
↑ Grzimek, 2003, с. 29
↑ Channing, Alan; Howell, Kim M. (2006). Amphibians of East Africa. Comstock Publishing. с. 104‑117. ISBN 0801443741.
↑ Sandberger, L.; Hillers, A.; Doumbia, J.; Loua, N-S.; Brede C.; Rödel, M-O. (2010). Rediscovery of the Liberian Nimba toad, Nimbaphrynoides liberiensis (Xavier, 1978) (Amphibia: Anura: Bufonidae), and reassessment of its taxonomic status. Zootaxa (Magnolia Press) 4355: 56‑68. ISSN 1175-5334.
↑ Gilbert, Perry W. (1942). Observations on the eggs of Ambystoma maculatum with especial reference to the green algae found within the egg envelopes. Ecology 23: 215‑227. JSTOR 1931088.
↑ Waldman, Bruce; Ryan, Michael J. (1983). Thermal advantages of communal egg mass deposition in wood frogs (Rana sylvatica). Journal of Herpetology 17 (1): 70‑72. JSTOR 1563783.
↑ Estrada, Alberto R.; Hedges, S. Blair (1996). At the lower size limit in tetrapods: a new diminutive frog from Cuba (Leptodactylidae: Eleutherodactylus). Copeia 1996 (4): 852‑859. JSTOR 1447647.
↑ Whittaker, Kellie; Chantasirivisal, Peera (2005-12-02). Leptodactylus pentadactylus. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-10-17. Процитовано 2012-07-19.
↑ Whittaker, Kellie (2007-27-06). Agalychnis callidryas. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-10-17. Процитовано 2012-07-19.
↑ Warkentin, K.M. (1995). Adaptive plasticity in hatching age: a response to predation risk trade-offs. Proceedings of the National Academy of Sciences 92 (8): 3507‑3510. doi:10.1073/pnas.92.8.3507.
↑ аб Stebbins, Robert C.; Cohen, Nathan W. (1995). A Natural History of Amphibians. Princeton University Press. с. 179‑194. ISBN 0-691-03281-5.
↑ Ontogenetic Changes in Diet and Intestinal Morphology in Semi-Terrestrial Tadpoles of Nannophrys ceylonensis (Dicroglossidae).
↑ Janzen, Peter (2005-05-10). Nannophrys ceylonensis. AmphibiaWeb. University of California, Berkeley. Архів оригіналу за 2012-10-17. Процитовано 2012-07-20.
↑ Duellman, W. E.; Zug, G. R. Anura: From tadpole to adult. Encyclopædia Britannica. Архів оригіналу за 2012-10-17. Процитовано 2012-07-13.
↑ Crump, Martha L. (1986). Cannibalism by younger tadpoles: another hazard of metamorphosis. Copeia (American Society of Ichthyologists and Herpetologists) 4: 1007‑1009. JSTOR 1445301.
↑ Stebbins, Robert C.; Cohen, Nathan W. (1995). A Natural History of Amphibians. Princeton University Press. с. 173‑175. ISBN 0-691-03281-5.
↑ Balinsky, Boris Ivan. Animal development: Metamorphosis. Encyclopedia Britannica. Архів оригіналу за 2012-10-17. Процитовано 2012-08-10.
↑ Campbell, Neil A.; Reece, Jane B. (2002). Biology (Sixth edition). Pearson Education. с. 692. ISBN 0-201-75054-6.
↑ Da Silva, H. R.; De Britto-Pereira, X. C. (2006). How much fruit do fruit-eating frogs eat? An investigation on the diet of Xenohyla truncata (Lissamphibia: Anura: Hylidae). Journal of Zoology 270 (4): 692–698. doi:10.1111/j.1469-7998.2006.00192.x.
↑ Graham, Donna. Northern leopard frog (Rana pipiens). An Educator's Guide to South Dakota's Natural Resources. Архів оригіналу за 2012-10-17. Процитовано 2012-08-04.
↑ Pimm, Stuart L. (1979). The structure of food webs. Theoretical Population Biology 16 (2): 144–158. PMID 538731. doi:10.1016/0040-5809(79)90010-8. Архів оригіналу за 2011-09-27.
↑ Matthews, K. R.; Miaud, C. (2007). A skeletochronological study of the age structure, growth, and longevity of the mountain yellow-legged frog, Rana muscosa, in the Sierra Nevada, California. Copeia 2007 (4): 986–993. ISSN 0045-8511. doi:10.1643/0045-8511(2007)7[986:ASSOTA]2.0.CO;2.
↑ Slavens, Frank; Slavens, Kate. Frog and toad index. Longevity. Архів оригіналу за 2012-10-17. Процитовано 2012-07-04.
↑ Crump, M. L. (1996). Parental care among the Amphibia. Advances in the Study of Behavior. Advances in the Study of Behavior 25: 109–144. ISBN 978-0-12-004525-9. doi:10.1016/S0065-3454(08)60331-9.
↑ абв Brown, J. L.; Morales, V.; Summers, K. (2010). A key ecological trait drove the evolution of biparental care and monogamy in an amphibian. American Naturalist 175 (4): 436—446. PMID 20180700. doi:10.1086/650727.
↑ Sheridan, Jennifer A.; Ocock, Joanne F. (2008). Parental care in Chiromantis hansenae (Anura: Rhacophoridae). Copeia 2008 (4): 733–736. doi:10.1643/CH-07-225.
↑ Brown, J. L.; Morales, V.; Summers, K. (2008a). Divergence in parental care, habitat selection and larval life history between two species of Peruvian poison frogs: An experimental analysis. Journal of Evolutionary Biology 21 (6): 1534—1543. PMID 18811668. doi:10.1111/j.1420-9101.2008.01609.x.
↑ Grant, T.; Frost, D. R.; Caldwell, J. P.; Gagliardo, R.; Haddad, C. F. B.; Kok, P. J. R.; Means, D. B.; Noonan, B. P.; Schargel, W. E.; Wheeler, W. (2006). Phylogenetic systematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae). Bulletin of the American Museum of Natural History 299: 1–262. ISSN 0003-0090. doi:10.1206/0003-0090(2006)299[1:PSODFA]2.0.CO;2.
↑ van Wijngaarden, René; Bolaños, Federico (1992). Parental care in Dendrobates granuliferus (Anura: Dendrobatidae), with a description of the tadpole. Journal of Herpetology 26 (1): 102–105. JSTOR 1565037. doi:10.2307/1565037.
↑ Fandiño, María Claudia; Lüddecke, Horst; Amézquita, Adolfo (1997). Vocalisation and larval transportation of male Colostethus subpunctatus (Anura: Dendrobatidae). Amphibia-Reptilia 18 (1): 39–48. doi:10.1163/156853897X00297.
↑ Common midwife toad, Alytes obstetricans. Repties et Amphibiens de France. Архів оригіналу за 2012-10-17. Процитовано 2012-07-30.
↑ Proteins of frog foam nests. University of Glasgow. Архів оригіналу за 2012-10-17. Процитовано 2012-08-24.
↑ Dalgetty, Laura; Kennedy, Malcolm W. (2010). Building a home from foam—túngara frog foam nest architecture and three-phase construction process. Biology Letters 6 (3): 293–296. PMC 2880057. PMID 20106853. doi:10.1098/rsbl.2009.0934.
↑ Assa darlingtoni. Frogs Australia Network. 2005. Процитовано 2012-08-05.
↑ Semeyn, E. (2002). Rheobatrachus silus. Animal Diversity Web. University of Michigan Museum of Zoology. Архів оригіналу за 2012-10-17. Процитовано 2012-08-05.
↑ Sandmeier, Fran (2001-03-12). Rhinoderma darwinii. AmphibiaWeb. Архів оригіналу за 2012-10-17. Процитовано 2012-08-05.
↑ Сербинова И. А. Размножение рогатой чесночницы (Megophrys nasuta) в Московском зоопарке // Научные исследования в зоологических парках : сборник. — 2010. — Вып. 26. — С. 22-31.
↑ Warkentin, I. G.; Bickford, D.; Sodhi, N. S.; Corey, J. A. (2009). Eating frogs to extinction. Conservation Biology 23 (4): 1056‑1059. doi:10.1111/j.1523-1739.2008.01165.x.
↑ Cultured Aquatic Species Information Programme: Rana catesbeiana. FAO: Fisheries and Aquaculture Department. Архів оригіналу за 2012-10-17. Процитовано 2012-07-05.
↑ Накормить Европу по-белорусски. Информационное агентство "Интерфакс-Запад". 04.03.2009. Архів оригіналу за 2012-10-18. Процитовано 2012-09-03.
↑ Wells, David Ames (1859). The science of common things: a familiar explanation of the first principles of physical science. For schools, families, and young students. Ivison, Phinney, Blakeman. с. 290.
↑ Stannius ligature. Biology online. 2005-10-03. Архів оригіналу за 2012-10-18. Процитовано 2012-08-05.
↑ Sarkar, S. (1996). Lancelot Hogben, 1895—1975. Genetics 142 (3): 655—660. PMC 1207007. PMID 8849876.
↑ Robert W. Briggs Biographical Memoir. Процитовано 2006-04-22.
↑ Browder, L.; Iten, L. (Eds.) (1998). Xenopus as a Model System in Developmental Biology. Dynamic Development. University of Calgary. Архів оригіналу за 2012-10-18. Процитовано 2012-06-17.
↑ Klein, S. Developing the potential of Xenopus tropicalis as a genetic model. Trans-NIH Xenopus Working Group. Архів оригіналу за 2012-10-18. Процитовано 2006-03-09.
↑ Xenopus tropicalis. Joint Genome Institute. Архів оригіналу за 2012-10-18. Процитовано 2012-07-13.
↑ Phillipe, G., Angenot L. (2005). «Recent developments in the field of arrow and dart poisons». J Ethnopharmacol 100(1-2): 85—91.
↑ VanCompernolle, S. E.; Taylor, R. J.; Oswald-Richter, K.; Jiang,J.; Youree, B. E.; Bowie, J. H.; Tyler, M. J.; Conlon, M.; Wade, D.; Aiken, C.; Dermody, T. S.; KewalRamani, V. N.; Rollins-Smith, L. A.; Unutmaz, D. (2005). Antimicrobial peptides from amphibian skin potently inhibit human immunodeficiency virus infection and transfer of virus from dendritic cells to T cells. Journal of Virology 79 (18): 11598–11606. PMC 1212620. PMID 16140737. doi:10.1128/JVI.79.18.11598-11606.2005.
↑ California Schools Leading Race to Stop Dissections. Animal Welfare Institute. 2011-04-25. Процитовано 2012-06-17.
↑ Lyttle, T.; Goldstein, D.; Gartz, J. (1996). Bufo toads and bufotenine: fact and fiction surrounding an alleged psychedelic. Journal of Psychoactive Drugs 28 (3): 267–290. PMID 8895112. doi:10.1080/02791072.1996.10472488.
↑ Lyttle, T. (1993). Misuse and legend in the "toad licking" phenomenon. International Journal of Addiction 28 (6): :521–538. PMID 8486435.
↑ Myers, Charles W.; Daly, John W.; Malkin, Borys (1978). A dangerously toxic new frog (Phyllobates) used by Emberá Indians of western Colombia, with discussion of blowgun fabrication and dart poisoning. Bulletin of the American Museum of Natural History 161: 307–366.
↑ Whittaker, Kellie; Lyser, Shelly (2005-02-14). Phyllobates terribilis. AmphibiaWeb. Архів оригіналу за 2012-10-18. Процитовано 2012-08-06.
↑ Phillipe, G.; Angenot L. (2005). Recent developments in the field of arrow and dart poisons. Journal of Ethnopharmacology 100 (1–2): 85–91. PMID 15993556. doi:10.1016/j.jep.2005.05.022.
↑ Stuart, S. N.; Chanson, J. S.; Cox, N. A.; Young, B. E.; Rodrigues, A. S. L.; Fischman, D. L.; Waller, R. W. (2004). Status and trends of amphibian declines and extinctions worldwide. Science 306 (5702): 1783‑1786. PMID 15486254. doi:10.1126/science.1103538.
↑ Voordouw, M. J.; Adama, D.; Houston, B.; Govindarajulu, P. (2010). Prevalence of the pathogenic chytrid fungus, Batrachochytrium dendrobatidis, in an endangered population of northern leopard frogs, Rana Pipiens. BMC Ecology 10 (6). doi:10.1186/1472-6785-10-6.
↑ Harp, Elizabeth M.; Petranka, James W. (2006). Ranavirus in wood frogs (Rana sylvatica): Potential sources of transmission within and between ponds. Journal of Wildlife Diseases 42 (2): 307‑318. PMID 16870853.
↑ Phillips, Kathryn (1994). Tracking the Vanishing Frogs. Penguin Books. ISBN 0-14-024646-0.
↑ Lips, Karen R. (2008). Decline of a tropical montane amphibian fauna. Conservation Biology 12: 106‑117. JSTOR 2387466. doi:10.1111/j.1523-1739.1998.96359.x.
↑ Blaustein, Andrew R.; Johnson, Pieter T. J. (2003). The complexity of deformed amphibians. Frontiers in Ecology and the Environment 1 (2): 87‑94.
↑ Burkhart, J. G.; Ankley, G.; Bell, H.; Carpenter, H.; Fort, D.; Gardiner, D.; Gardner, H.; Hale, R.; Helgen, J. C.; Jepson, P.; Johnson, D.; Lannoo, M.; Lee, D.; Lary, J.; Levey, R.; Magner, J.; Meteyer, C.; Shelby, M. D.; Lucier, G. (2000). Strategies for assessing the implications of malformed frogs for environmental health. Environmental Health Perspectives 108 (1): 83: 83‑90. JSTOR 3454299. PMC 1637865. PMID 10620528. doi:10.2307/3454299.
↑ Frog population decrease mostly due to traffic. New Scientist. 2006-07-07. Процитовано 2012-07-13.
↑ Black, Richard (2005-10-02). New frog centre for London Zoo. BBC News. Процитовано 2008-11-03.
↑ National recovery plan for the Southern Corroboree Frog (Pseudophryne corroboree): 5. Previous recovery actions. Environment.gov.au. Архів оригіналу за 2012-10-18. Процитовано 2008-11-03.
↑ Bacteria show promise in fending off global amphibian killer. Physorg.com. 2007-05-23. Архів оригіналу за 2012-10-18. Процитовано 2008-11-03.
↑ Project launched to fight frog-killing fungus. The Guardian. 2009-05-12. Процитовано 2009-07-19.
↑ 2008: Year of the Frog. 2008-01-15. Архів оригіналу за 2012-10-18. Процитовано 2012-07-13.
↑ Борейко В. Е. Биоксенофобия народных воззрений в отношении диких животных и растений (російською). Киевский эколого-культурный центр. Архів оригіналу за 2012-10-18. Процитовано 2012-08-27.
↑ Cooper, J.C. (1992). Symbolic and Mythological Animals. London: Aquarian Press. с. 106–08. ISBN 1-85538-118-4.
↑ Burns, William E. (2003). Witch Hunts in Europe and America: An Encyclopedia. Greenwood Publishing Group. с. 7. ISBN 0-313-32142-6.
↑ Berrin, Katherine; Larco Museum (1997). The Spirit of Ancient Peru: Treasures from the Museo Arqueológico Rafael Larco Herrera. New York: Thames and Hudson. ISBN 0-500-01802-2.
↑ Gratwicke, B. (2009). The Panamanian Golden Frog. Panama Amphibian Rescue and Conservation Project. Архів оригіналу за 2012-10-18.
↑ Жайворонок В. В. Знаки української етнокультури: Словник-довідник. — К.: Довіра, 2006. — 706 с. — С. 215.
↑ Grzimek, 2003, с. 52
↑ Культи, тварини, птахи / Завадська В., Музиченко Я. та ін. 100 найвідоміших образів української міфології. — К.: Орфей, 2002. — 448 с.
ISBN 966-96200-0-7
↑ Brothers Grimm. The Frog Prince. East of the Web. Архів оригіналу за 2012-10-18. Процитовано 2012-06-30.
Література |
Вікіцитати містять висловлювання на тему: Безхвості |
| Вікісховище має мультимедійні дані за темою: Безхвості |
Beltz, Ellin (2005). Frogs: Inside their Remarkable World. Firefly Books. ISBN 1-55297-869-9.
Cogger, H.G.; R.G. Zweifel, and D. Kirschner (2004). Encyclopedia of Reptiles & Amphibians Second Edition. Fog City Press. ISBN 1-877019-69-0.- Estes, R., and O. A. Reig. (1973). «The early fossil record of frogs: a review of the evidence.» pp. 11–63 In J. L. Vial (Ed.), Evolutionary Biology of the Anurans: Contemporary Research on Major Problems. University of Missouri Press, Columbia.
Gissi, Carmela; Diego San Mauro, Graziano Pesole and Rafael Zardoya (February 2006). Mitochondrial phylogeny of Anura (Amphibia): A case study of congruent phylogenetic reconstruction using amino acid and nucleotide characters. Gene 366 (2): 228—237. PMID 16307849. doi:10.1016/j.gene.2005.07.034.
Holman, J. A (2004). Fossil Frogs and Toads of North America. Indiana University Press. ISBN 0-253-34280-5.
Tyler, M. J. (1994). Australian Frogs A Natural History. Reed Books. ISBN 0-7301-0468-0.
Grzimek, B. (2003). Grzimek’s Animal Life Encyclopedia. 6, Amphibians (вид. 2nd edition). Farmington Hills: Gale Group. с. 3—321. ISBN 0-7876-5362-4. Процитовано October 7, 2012.
Українська радянська енциклопедія : у 12 т. / гол. ред. М. П. Бажан ; редкол.: О. К. Антонов та ін. — 2-ге вид. — К. : Головна редакція УРЕ, 1974–1985.- Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М.: Сов. энциклопедия, 1986. — 831 с. (рос.)